
Allosaurus
| Evolúciós időszak: késő jura, 155–150 Ma | ||||||||||||||||||||
 Allosaurus modell Lengyelországban
| ||||||||||||||||||||
| Természetvédelmi státusz | ||||||||||||||||||||
| Fosszilis | ||||||||||||||||||||
| Rendszertani besorolás | ||||||||||||||||||||
| ||||||||||||||||||||
| Szinonimák | ||||||||||||||||||||
| ||||||||||||||||||||
| Fajok | ||||||||||||||||||||
| Elterjedés | ||||||||||||||||||||

| ||||||||||||||||||||
| Hivatkozások | ||||||||||||||||||||
A Wikifajok tartalmaz Allosaurus témájú rendszertani információt. A Wikimédia Commons tartalmaz Allosaurus témájú médiaállományokat és Allosaurus témájú kategóriát. |
Az Allosaurus (jelentése 'különös gyík', az ógörög αλλος / allosz 'furcsa', 'eltérő' és σαυρος / szaürosz 'gyík' szavak összetételéből)[1] a nagy theropoda dinoszauruszok egy neme. Fajai főként a jura időszakban, mintegy 155-150 millió évvel ezelőtt éltek a mai Észak-Amerikában. Az első, egyértelműen ehhez a nemhez tartozó maradványokat Othniel Charles Marsh írta le 1877-ben. Mivel ez az első jól ismert theropoda dinoszaurusz, nem csak az őslénykutatók figyeltek fel rá, több játék- illetve dokumentumfilm is foglalkozott vele.
Az Allosaurus két lábon járó húsevő volt, nagy koponyájában tucatnyi éles foggal. Testhossza körülbelül 8,5 méter lehetett, de a töredékek alapján elképzelhető, hogy akadtak 12 méteres példányok is. Hátsó lábai aránylag nagyok és erőteljesek; mellső lábain három ujj nőtt. Testét a hosszú és nehéz farok tartotta egyensúlyban. Allosauridaként, a Carnosauria theropodák egy csoportjaként osztályozták. A nem taxonómiája bonyolult: legismertebb faja az A. fragilis, de számos tisztázatlan érvényű fajt is ide sorolnak. Maradványainak többsége az észak-amerikai Morrison-formációból került elő, de ismertek portugál és feltételesen ide sorolt tanzániai leletek is. A 20. század közepéig Antrodemus néven ismerték, de a Cleveland-Lloyd dinoszaurusz-lelőhelyről (Cleveland-Lloyd Dinosaur Quarry) előkerült bőséges maradványok hatására megkapta az Allosaurus nevet, és az egyik legismertebb dinoszaurusszá vált.
A Morrison-formáció egyik jelentős nagy méretű ragadozójaként a tápláléklánc csúcsán helyezkedhetett el, és elképzelhető, hogy a korabeli nagy növényevőkre vadászott, de talán más húsevőkre (például a Ceratosaurusra) is. Potenciális zsákmányai közé tartoztak az ornithopodák, a stegosauridák és a sauropodák. Bár gyakran úgy gondolják, hogy a sauropodákra csoportosan vadászott, kevés bizonyíték áll rendelkezésre az együttműködő szociális viselkedésére vonatkozóan, és az is elképzelhető, hogy az egyes példányok inkább agresszíven viselkedtek egymással, így az egy csoportban összegyűlt maradványok talán egy zsákmányból közösen táplálkozó magányos egyedek lehettek. Feltételezhető, hogy lesből csaptak le az áldozataikra, szekerceként használva a felső állcsontjukat.
Anatómia[szerkesztés]


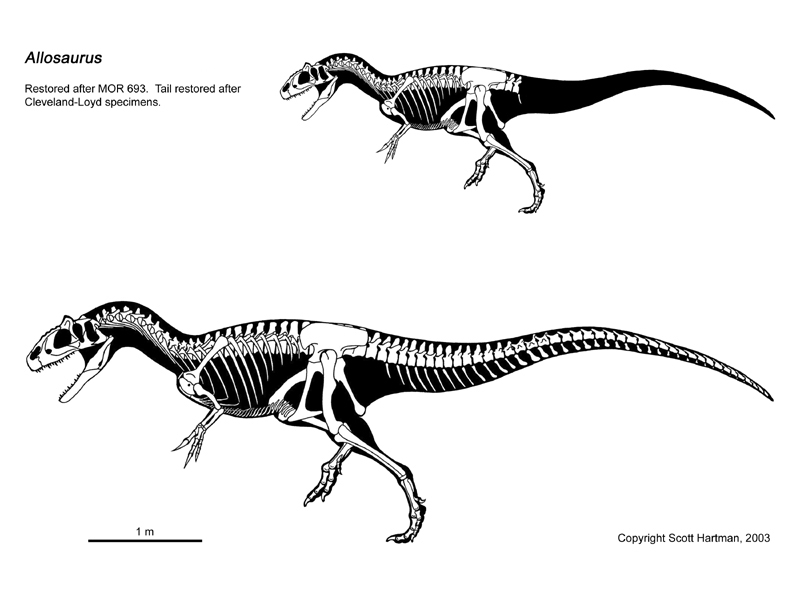
Az Allosaurus egy szokványos nagyméretű theropoda, rövid nyakon ülő masszív koponyával, hosszú farokkal és kisméretű mellső lábakkal rendelkezett. Legismertebb faja, az Allosaurus fragilis átlagos hossza 8,5 méter,[2] a legnagyobb példányának (az Amerikai Természetrajzi Múzeum AMNH 680-as jelzésű leletének) becsült hosszúsága 9,7 méter,[3] a tömege pedig mintegy 2,3 tonna lehetett.[3] 1976-ban James Madsen az Allosaurusról készült írásában a csontok mérete alapján a maximális hosszúságot 12–13 méterre becsülte.[4] Ahogy a dinoszauruszoknál általában, a tömegbecslés viták tárgyát képezi, és 1980 óta a felnőtt egyedekre vonatkozóan 1500 kilogrammtól 1000–4000 kilogrammon át 1010 kilogrammig többféle átlagos tömeget valószínűsítettek.[5] A Morrison-formáció szakértője, John Foster kijelentette, hogy az 1000 kilogramm megfelel egy nagyméretű, felnőtt A. fragilisnek, míg a 700 kilogramm közelítő becslés az általa mért átlagos combcsontú egyedek számára.[6]
Több óriási példányt is Allosaurusnak véltek, de elképzelhető, hogy más nemhez tartoztak. A közeli rokonságába tartozó Saurophaganax (a Sam Noble Oklahomai Természetrajzi Múzeum OMNH 1708-as lelete) elérhette a 10,9 méteres testhosszt,[3] és egyes példányait időnként Allosaurus maximusként sorolták be, de az újabb keletű vizsgálatok szerint más nemhez tartoznak.[7] Egy másik feltételezett Allosaurus példányt (az AMNH 5767 jelzésűt), amit egyszer az Epanterias nemhez soroltak be, 12,1 méter hosszúságúra becsültek.[3] Az egyik legújabb felfedezés egy Peterson-lelőhelyről, a Morrison-formáció Új-Mexikó állambeli részéről származó hiányos csontváz; ez a nagy méretű allosaurida feltehetően egy újabb Saurophaganax példány.[8]
Koponya[szerkesztés]

Az Allosaurus koponyája és fogazata mérsékelten arányul a theropoda méreteihez. Gregory S. Paul őslénykutató egy 845 milliméter hosszú koponyához 7,9 méteres testhosszt valószínűsített.[9] Az egyes premaxillák (a pofa elejét alkotó csontok), öt darab D keresztmetszetű fogat tartanak, a maxillákban (a felső állcsont fő fogtartó csontjaiban) pedig 14–17 fog helyezkedik el; a fogak száma nem függ össze pontosan a csont méretével. Az alsó állkapocs fogmederként szolgáló (dentális) csontjai 14–17, átlagosan 16 fogat tartanak. A fogak rövidebbek, keskenyebbek és nagyobb ívben hajlanak hátra a koponya hátsó része közelében. Az összes fog fűrészszerű éllel rendelkezik. Könnyen kihullottak, de folyamatosan pótlódtak, a fosszíliákban gyakran megtalálhatók.[4]

A koponya legjellegzetesebb részei a szemek felett lévő csontképződmények. Ezek a szarvak, melyek a könnycsont kiterjedései,[4] különböző alakúak és méretűek. Az orrcsontok felső éleitől dupla redők futnak a szarvakig.[4] A szarvak, melyeket valószínűleg keratin burkolat fedett, talán többféle célt is szolgáltak, például védték a szemet a napfénytől,[4] pózolásnál vagy a fajtársakkal vívott harcokban lehetett szerepük[9][10] (habár törékenyek voltak).[4] A koponyatető hátsó részén egy, a tyrannosauridáknál is megtalálható redő helyezkedik el, ami izomtapadási pontként szolgált.[9]
A könnycsontok belsejében levő benyomódásokban talán mirigyek (például sómirigyek) lehettek.[11] A maxillákban üregek voltak, valamivel fejlettebbek azoknál, amik a bazális theropodáknál, például a Ceratosaurusnál és a Marshosaurusnál ismertek; és feltehetően a szaglásban volt szerepük, talán a vomeronazális szervhez (Jacobson-szerv) hasonlót tartalmaztak. A agyat fedő koponyatető vékony volt, feltehetően az agy hőszabályzásának javítására.[4] A koponya és az állkapocscsont ízületei lehetővé tették ezen részek elmozdulását. Az állkapocs első és hátsó fele lazán illeszkedett, miáltal ez a testrész kifelé tudott hajolni, így az állat nagyobbra tudta nyitni a száját.[12] A koponyatető és a homlokrész talán szintén ízülettel volt ellátva.[4]
A csontváz egyéb részei[szerkesztés]

Az Allosaurus 9 nyak-, 25 hát-, és 5 keresztcsonti (medence) csigolyája volt[4] (a nyak és a hát kezdetét illetően több eltérő álláspont is létezik, például Gregory S. Paul szerint a nyak 10, a hát pedig 13 csigolyából áll). A farokcsigolyák száma ismeretlen, illetve egyedenként eltérő; James Madsen 50-re becsüli a számukat,[4] míg Gregory S. Paul szerint a farok mindössze 45 vagy kevesebb csigolyából állt.[9] A nyaki és elülső hátcsigolyákban üreges részek voltak.[4] Hasonló képződmények találhatók a modern theropodákban (és a madarakban is), melyekről azt gondolják, hogy a légzőrendszer részét képező légzsákokat tartalmaztak.[13] A bordák terjedelmesek voltak, különösen az olyan kevésbé fejlett theropodákhoz viszonyítva, mint a Ceratosaurus,[14] és hordó alakú mellkast képeztek. Az Allosaurus rendelkezett hasi bordákkal (gastralia) is, de ezek ritkán kerülnek elő,[4] és talán a többinél kevésbé lehettek csontosak.[9] Egy publikált esetnél a hasi bordák az állat élete során bekövetkezett sérülés jelét mutatták.[15] A villacsont szintén a váz részét képezte, de csak 1996-ban ismerték fel; egyes esetekben ugyanis a villacsontot összetévesztették a hasi bordákkal.[15][16] A csípőcsont, a csípő fő része erőteljes, a szeméremcsonton pedig egy kiemelkedő alsó rész helyezkedett el, ami egyfelől izomtapadási pontként szolgálhatott, másfelől pedig megtámaszthatta a testet a földön, nyugvó helyzetben. Madsen feljegyezte, hogy a Cleveland-Lloyd dinoszaurusz-lelőhelyről előkerült példányok mintegy felénél, a mérettől függetlenül előfordul, hogy a szeméremcsontok nem nőttek össze egymással az alsó résznél. Véleménye szerint ez a nemi kétalakúság (ivari dimorfizmus) jele, és a nőstényeknél az összeforratlan csontok a tojások kijutását segítik.[4] Elképzelése azonban nem váltott ki tudományos érdeklődést.

A mellső lábak jóval rövidebbek voltak a hátsóknál (a felnőtteknél csak a hátsó láb hosszának 35%-át érik el),[17] a rajtuk levő három ujj nagy, erősen görbült és hegyes karmokkal volt ellátva,[4] melyek hossza elérte a 15 centimétert is.[18] A mellső lábak erőteljesek voltak,[9] és az alkar valamivel rövidebb volt a felkarnál (a felkarcsont/singcsont aránya 1:1,2).[19] A csukló egy félholdas kéztőcsonttal[20] rendelkezett, ami más fejlettebb theropodáknál, például a maniraptoráknál is megtalálható. A három ujj közül a legbelső (a hüvelykujj) volt a legnagyobb,[9] és eltért a többitől, az állat befelé tudta fordítani. A hátsó lábak nem voltak olyan hosszúak vagy sebes mozgáshoz alkalmazkodottak, mint a tyrannosauridák esetében, a karmaik pedig a korábbi theropodákéhoz hasonlítva kevésbé fejlettek, inkább pataszerűnek[9] tűnnek. Minden lábon három, nagy teherbírású karom volt és egy belső fűkarom, ami Madsen szerint a fiókák megragadására szolgált.[4] Az ötödik (legszélső) lábközépcsonthoz egy szárkapocscsontszerű maradvány tartozott, ami talán emelőként szolgált az achilles-ín és a lábfej között.[21] Madsen 1976-ban ezeket a csontokat a belső lábközépcsont lehetséges felső részeiként azonosította.
Osztályozás[szerkesztés]
Az Allosaurus egy allosaurida volt, a nagy theropodák családjába tartozó Carnosauria csoport tagja. Az Allosauridae családot 1878-ban Othniel Charles Marsh hozta létre a nem számára,[22] de az az 1970-es évekig feledésbe merült, mert a kutatók inkább a szemétkosár-taxonként használt Megalosauridae-t részesítették előnyben. Ekkortájt hagytak fel az Allosaurus Antrodemusként való megnevezésével is, de a régi név továbbra is fontos lehet azok számára, akik James Madsen 1976-os művénél korábbi publikációkban keresnek információkat az Allosaurusra vonatkozóan. A Megalosauridae családra hivatkozó hosszabb írások között Charles Gilmore (1920),[19] Friedrich von Huene (1926),[23] Alfred Sherwood Romer (1956, 1966),[24][25] R. Steel (1970)[26] és Alick Walker (1964)[27] munkája is megtalálható.
Madsen művének hatására az Allosauridae elfogadott családdá vált, a definíciója azonban nem volt megfelelő. A félig szakmai jellegű munkák a családot különböző nagy testű theropodákhoz társították, főként azokhoz, amelyek nagyobbak és ismertebbek voltak a megalosauridáknál. Az Allosaurus rokonának tekintett theropodák között megtalálható az Indosaurus, a Piatnitzkysaurus, a Piveteausaurus, a Yangchuanosaurus,[28] az Acrocanthosaurus, a Chilantaisaurus, a Compsosuchus, a Stokesosaurus és a Szechuanosaurus[29] is. A theropodák sokféleségével kapcsolatos modern ismeretek és az evolúciós kapcsolatok kladisztikus vizsgálatainak hatására ezeket a theropodákat már nem tekintik allosauridának, néhányuk azonban, például az Acrocanthosaurus és a Yangchuanosaurus a rokon családok tagjai.[13]
Az Allosauridae egyike a három Carnosauria családnak; a másik kettő a Carcharodontosauridae és a Sinraptoridae.[13] Az Allosauridae-t időnként a Tyrannosauridae ősének tekintették (miáltal parafiletikus csoporttá vált), erre az egyik legújabb példa Gregory S. Paul Predatory Dinosaurs of the World (A világ ragadozó dinoszauruszai)[30] című műve, ám ezt a nézetet elvetették, amikor a tyrannosauridákat a Coelurosauria csoportba, a theropodák egy külön ágaként sorolták be.[31] Az Allosauridae a legkisebb carnosaurida család, az Allosaurus mellett csak a Saurophaganax és egy névtelen, francia allosauroidea tartozik az elfogadott nemei közé.[13] Lehetséges, hogy az Epanterias is ide sorolható, de az sem kizárt, hogy a Saurophaganax mellett ez a nem is csak az Allosaurus egy nagy méretű példánya.[9] A legújabb ismertetők megtartják a Saurophaganax nemet, az Epanteriast pedig az Allosaurus szinonimájának tekintik.[5][13]
Felfedezések[szerkesztés]
Korai felfedezések és kutatások[szerkesztés]
Az Allosaurus felfedezése és korai vizsgálatai nehezen áttekinthetőek, mivel az 1800-as években lezajlott „csontháború” idején sokféle elnevezést használtak. Az első fosszíliáról készült leírás 1869-ből, Ferdinand Vandiveer Haydentől származik, aki másodkézből jutott egy csonthoz, ami feltehetően a Middle Parkból, a Morrison-formáció területéről, Granby közeléből származott. A helybéliek a hasonló csontokat „megkövült ló pataként” azonosították. Hayden elküldte a leletet Joseph Leidynek, aki megállapította, hogy egy farokcsigolya fele, és átmenetileg az európai Poekilopleuron nem Poicilopleuron [sic] valens nevű fajaként sorolta be.[32] Később úgy döntött, hogy saját nemet alkot a számára, az Antrodemust.[33]
Az Allosaurus holotípusa, az YPM 1930 egy töredékes csontokból álló gyűjtemény, ami tartalmaz három csigolyát, egy borda-maradványt, egy fogat, egy lábcsontot és a jobb felkarcsont szárát, ami a legtöbb vita alapjául szolgál. Othniel Charles Marsh 1877-ben az Allosaurus fragilis nevet adta a leletnek. Az Allosaurus név az órörög αλλος / allosz ('furcsa', 'eltérő') és σαυρος / szaürosz ('gyík') szavak összetételéből származik.[1] Azért nevezték el „különös gyíknak”, mert a gerince eltért azokétól a dinoszauruszokétól, amiket a felfedezése idején ismertek.[34][35] A faj jelzője, a fragilis (a latin szó jelentése: törékeny), a könnyített csigolyákra utal. A kövületeket a Colorado állambeli Cańon Citytől északra, a Garden Park területén, a Morrison-formációban találták meg.[34] Marsh és vetélytársa, Edward Drinker Cope számos különböző nemet azonosított a későbbiekben az Allosaurusénak tulajdonított szórványos anyagokhoz való hasonlóság alapján. Ezek között volt a Marsh által felfedezett Creosaurus[22] és a Labrosaurus,[36] valamint a Cope által elnevezett Epanterias.[37]


Sietségükben Cope és Marsh nem mindig ellenőrizték a felfedezéseiket (vagy általában inkább az alárendeltjeikre bízták ezt a feladatot). Például Benjamin Franklin Mudge-ot, az Allosaurus típusfajának coloradoi felfedezőjét Marsh egy wyomingi feladatra jelölte ki 1883-ban, amikor folytatódott a munka a Garden Parkban, ahol M. P. Felch talált egy majdnem teljes Allosaurust és több részleges csontvázat.[11] Továbbá Cope egyik begyűjtője, H. F. Hubbell 1879-ben a wyomingi Como Bluff területén talált egy példányt, de mivel nem számolt be a teljességéről, Cope sosem csomagolta ki. Csak 1903-ban vizsgálták meg (évekkel Cope halála után), és kiderült, hogy az egyik addig ismert legteljesebb theropoda példány. 1908-ban AMNH 5753-as jelzéssel katalogizálták, és bemutatták a nyilvánosság előtt.[38] Később Charles R. Knight illusztrációt készített róla, amelyen a jól ismert helyzetben, egy részleges Apatosaurus fölé tornyosulva áll, látszólag dögevőként táplálkozva. Habár az első szabadon felállított theropoda dinoszauruszként vált ismertté, melyet gyakran használnak illusztrációként és fényképeznek, mégsem készült róla tudományos leírás.[39]

A korai nevek sokfélesége megnehezítette a későbbi kutatásokat, ahogy az is, hogy Mash és Cope mindössze rövid leírásokat készített. Ekkoriban Samuel Wendell Williston és más kutatók felvetették, hogy túl sok név van használatban.[40] Például 1901-ben Williston rámutatott, hogy Marsh sosem volt képes megfelelő módon különbséget tenni az Allosaurus és a Creosaurus között.[41] 1920-ban Charles W. Gilmore volt a legbefolyásosabb azok közül, akik megpróbálták rendezni a kaotikus helyzetet. Arra a következtetésre jutott, hogy a farokcsigolya, amit Leidy az Antrodemushoz társított, megkülönböztethetetlen az Allosaurusétól, így az Antrodemusnak kell elfogadott névvé válnia, mivel régebbiként elsőbbséget élvez.[19] Ezután ötven évre az Antrodemus lett a nem elfogadott neve, míg végül James Madsen publikálta a Cleveland-Lloyd példányokat, és úgy döntött, hogy az Allosaurus nevet fogja használni, mivel az Antrodemus gyenge tárgyi bizonyítékokon, diagnosztikus jellemzőkön és lelőhely információn alapul, már amennyire vannak ilyenek (nem ismert ugyanis, hogy az Antrodemus egyetlen csontja melyik formációból származik).[4] Az „Antrodemus” elnevezést nem hivatalosan arra használják, hogy különbséget tegyenek a Gilmore által rekonstruált és a Madsen által összeállított és rekonstruált koponya között.[42]
A Cleveland-Lloyd lelőhely[szerkesztés]
A szórványos munkákból már 1927-ben ismertté vált a Utah állambeli Emery Countyban levő Cleveland-Lloyd dinoszaurusz-lelőhely, de fosszília lelőhelyként William J. Stokes írt róla elsőként 1945-ben,[43] nagyobb munkálatok pedig nem folytak a területen az 1960-as évek előtt. 1960 és 1965 között, közel 40 intézmény összefogása révén több ezer csontot tártak fel.[4] A lelőhely nevezetes a jelentős mennyiségű Allosaurus maradványról, az ott található egyedek jó állapotáról, és arról, hogy nincs részletes tudományos magyarázat arra, hogyan jött létre. A csontok többsége a nagy testű theropodához, az Allosaurus fragilishez tartozik (a becslések alapján az ott talált minimum 73 dinoszauruszból mintegy 46 A fragilis). A fosszíliák széttagolódva és összekeveredve helyezkednek el. Közel egy tucatnyi tudományos cikket jelentettek meg a lelőhely tafonómiájáról, melyek számos egymásnak ellentmondó magyarázatot tartalmaznak a hely kialakulására vonatkozóan. Az elképzelések között például szerepel az, hogy az állatok mocsárba vagy mély sárba ragadva pusztultak el, hogy az aszály idején egy vízzel teli mélyedésbe fulladtak bele, illetve hogy egy tavaszi áradás végzett velük egy tó közelében vagy egy ártéri területen.[44] Eltekintve a magyarázatoktól, a nagy számú, jól megőrződött Allosaurus maradvány lehetővé tette a nem részletes megismerését, miáltal bekerült a legjobban ismert theropodák közé. A lelőhely csontmaradványai különféle korú és méretű egyedekhez tartoznak, melyek hossza 1 métertől[45] 12 méterig változik, és a széttagoltságuk előnyt jelent a rendszerint csak összeforrva megtalálható csontok leírásánál.[4]
Újabb felfedezések: 1980-tól napjainkig[szerkesztés]
Azt az időszakot, amely Madsen művének megjelenése óta eltelt, kiterjedt kutatások fémjelezték az Allosaurus életével kapcsolatos (ősbiológiai és ősökológiai) témaköröket illetően. A vizsgált témák között szerepel a csontok különbözősége,[46] a növekedés,[47][48] a koponya szerkezete,[49] a vadászati módszerek,[50] az agy működése[51] és az esetleges társas életmód, illetve a szülői gondoskodás.[52] A régi kutatási anyagok újraelemzése (főként a nagy példányokra vonatkozóan),[9][53] az új portugáliai felfedezések[54] és a különböző majdnem teljes új példányok[15][55][56] tovább bővítették a tudományos ismereteket. Gerard Gierliński lengyel őslénykutató a 2000-es évek elején egy sor Bałtówban talált megkövesedett lábnyomot az Allosaurusénak tulajdonított.[57]
„Nagy Al”[szerkesztés]

Az egyik legjelentősebb Allosarurus lelet (A Museum of the Rockies MOR 693-as jelzésű példánya), az 1991-ben felfedezett „Nagy Al” néven ismertté vált, 95%-ban teljes, részben tagolt csontváz, ami 8 méter hosszú. A wyomingi Shell közelében bukkant rá egy svájci kutatócsoport, melyet Kirby Siber vezetett, a feltárást pedig a Museum of the Rockies és a Wyomingi Egyetem Geológiai Múzeuma (University of Wyoming Geological Museum) tudósai együtt végezték el.[58] A svájci csoport később kiásott egy újabb Allosaurust, amely az eddigi legjobb állapotban megőrződött fosszília, és a „Nagy Al 2” nevet kapta.[56] A „Nagy Al” név a lelet teljességét, jó állapotát és tudományos fontosságát jelzi; maga a példány egy közepesnél kisebb méretű,[58] nagyjából 87%-ban kifejlett állat volt.[59] A fosszíliáról Brent H. Breithaupt készített leírást 1996-ban.[55] Csontjai közül 19 eltört, illetve betegség jeleit mutatja, ami valószínűleg közrejátszott az állat pusztulásában. A beteg csontok közé öt borda, öt csigolya és négy lábcsont tartozik; egy részükön a csont fertőzése, csontvelőgyulladás nyoma látható. A legnagyobb problémát az élő állat számára az a fertőzés és trauma jelenthette, ami a jobb lábát érintette, és valószínűleg kihathatott a mozgására, illetve a járása megváltozása révén talán fogékonyabbá tehette egyéb lábsérülésekre is.[59]
Fajok és taxonómia[szerkesztés]

Tisztázatlan hogy az Allosaurusnak hány faja volt. 1988-ig hét érvényes fajt tartottak számon (az A. amplexus,[9] az A. atrox,[9] az A. europaeus,[60] a típusfaj az A. fragilis,[13] a hivatalosan le nem írt „A. jimmadseni”,[7] az A. maximus[46] és az A. tendagurensis[13]), azonban rendszerint egyszerre csak egy töredéket tekintenek érvényesnek. Emellett legalább tíz bizonytalan vagy leíratlan faj létezik, melyet az évek során az Allosaurus nemhez társítottak, azon nemek mellett, amik a későbbiekben beolvadtak az Allosaurusba. A bazális tetanurán theropodákról szóló legújabb ismertető szerint csak az A. fragilis (az A. amplexust és az A. atroxot szinonimájaként véve), az „A. jimmadseni” (mint névtelen faj), és az A. tendagurensis az elfogadott és érvényes fajok, míg az A. europaeus egyelőre nem számít annak, az A. maximus pedig a Saurophaganax nemhez tartozik.[13]
Az A. amplexus, az A. atrox, az A. fragilis, az „A. jimmadseni” és az A. maximus maradványait mind az Amerikai Egyesült Államokban levő Morrison-formációban, Colorado, Montana, Új-Mexikó, Oklahoma, Dél-Dakota, Utah és Wyoming államok területén elszórtan, a késő jura kor kimmeridge-i–tithon korszakához tartozó rétegekben találták meg. Az A. fragilis a leggyakoribb, legalább 60 példány alapján ismert faj.[13] Az 1980-as évek óta viták folynak arról, hogy a Morrison-formációban talán két Allosaurus faj élt, melyek közül a második az A atrox lehetett;[9][61] az újkeletű művekben azonban továbbra is az „egy faj” elképzelés jelenik meg,[13] a Morrison-formációból előkerült leletek eltéréseit csak az egyedek közti különbségeknek tekintik.[62][63] A Cleveland Lloyd dinoszaurusz-lelőhely példányai kisebbek és könnyebb felépítésűek, mint az a hatalmas és robusztus egyed, amely a Dry Mesa lelőhelyről került a Brigham Young Egyetem birtokába, de a különböző lelőhelyeken talált csontok alakján nem látható eltérés.[62] Az A. europaeusra a Lourinhă-formáció részét képező Porto Novo-tagozat kimmeridge-i korszakhoz tartozó rétegeiben bukkantak rá,[60] de feltételezhető, hogy a lelet egy A. fragilis.[64] Az A. tendagurensis a tanzániai Mtwara régióban levő Tendaguru kimmeridge-i korszakához tartozó rétegeiből került elő.[65] Habár a legújabb ismertetők átmenetileg érvényes fajnak fogadják el, elképzelhető, hogy a maradványok egy bazálisabb tetanuránhoz[66] vagy egy nem meghatározható (nomen dubium – kétséges nevű) theropodához tartoznak.[2] Annyi bizonyos, hogy nagy termetű, körülbelül 10 méter hosszú és nagyjából 2,5 tonna tömegű állat lehetett.[3]
Az Allosaurust az Antrodemus, a Creosaurus, az Epanterias, valamint a Labrosaurus nem szinonimájának tekintik.[13] A legtöbb, törmelékes maradványok alapján ismert fajt az A. fragilis szinonimájának tekintik vagy nem társítják ehhez a nemhez. Kivételt képez az 1884-ben Marsh által elnevezett Labrosaurus ferox, amit egy különös formájú állkapocs alapján azonosított, aminek a hegyénél a fogsoron egy feltűnő lyuk látható, és amelynek hátsó része nagy mértékben nyújtott és lefelé irányul.[67] A későbbiekben a kutatók felvetették, hogy a csont kóros elváltozás folytán deformálódott, és az élő állat sérülését jelzi,[19] a hátsó rész szokatlan formája pedig a gipsszel való rekonstrukció eredménye.[68] Ezt a leletet jelenleg az A. fragilis példányának tekintik.[13] A világ számos pontján találtak olyan fosszíliákat, amelyeket az Allosaurusénak tulajdonítanak, például Ausztráliában,[69] Szibériában[70] és Svájcban,[2] de ezekről megállapították, hogy más dinoszauruszokhoz tartoznak.
Ősökológia[szerkesztés]

Az Allosaurus a leggyakoribb nagy testű theropoda volt a mai Egyesült Államok nyugati részén, mint ahogy arról a Morrison-formáció szikláiban megőrződött kövületek tanúskodnak (a theropodák 70–75%-át teszi ki,[6]) és ezen a területen nyilván a tápláléklánc csúcsán állt.[71] A Morrison-formáció félig száraz éghajlatú környezet lehetett, nedves és száraz évszakokkal, és egy lapos ártéri területtel.[72] A növényzet az ártéri erdőktől, a tűlevelűekkel, páfrányfákkal és harasztokkal borított térségeken át a fákkal ritkán benőtt szavannás területekig meglehetősen változékony volt.[73]
A Morrison-formáció gazdag terület a fosszíliavadászok számára, zöld algák, gombák, mohák, zsurlók, páfrányok, cikászok, ginkók és a tűlevelűek számos családjának maradványai találhatók itt. Az egyéb fosszíliák között előfordulnak kétkagylósok, csigák, sugarasúszójú halak, békák, szalamandrák, teknősök, sphenodonták, gyíkok, szárazföldi és vízi crocodylomorphák, a pterosaurusok különböző fajai, több dinoszauruszfaj, valamint olyan korai emlősök, mint a docodonták, a multituberculaták, a symmetrodonták és a triconodonták. A dinoszauruszok között előfordulnak theropodák, mint például a Ceratosaurus, az Ornitholestes és a Torvosaurus, sauropodák, mint például az Apatosaurus, a Brachiosaurus, a Camarasaurus és a Diplodocus, valamint madármedencéjűek, mint a Camptosaurus, a Dryosaurus és a Stegosaurus.[74] Portugália késő jura kori formációi, melyek területén az Allosaurus is élt, hasonlítanak a Morrisonra, de erős óceáni hatás érezhető rajtuk. A két terület számos dinoszaurusza ugyanahhoz a nemhez (például Allosaurus, Ceratosaurus, Torvosaurus, és Apatosaurus) vagy hasonlóhoz (például Brachiosaurus, Lusotitan, Camptosaurus és Draconyx) tartozott.[75]

Az Allosaurus mellett mindkét területen olyan nagy testű theropodák éltek, mint a Ceratosaurus és a Torvosaurus.[75] A fosszíliák lelőhelye és anatómiai vizsgálata alapján mindhárom különböző ökológiai fülkét töltött be. A ceratosaurusok és a torvosaurusok feltehetően a vízi utak mentén éltek, ahol vékonyabb testük előnyt jelenthetett az erdős és aljnövényzettel benőtt területeken, míg az Allosaurusok zömök teste és hosszú lábai gyorsabb mozgást, de kevésbé jó manőverező képességet biztosítottak, és úgy tűnik, inkább a kiszáradt ártéri területeket részesítették előnyben.[76] A Torvosaurusnál jobban ismert Ceratosaurust magasabb és keskenyebb koponyája, valamint széles fogai teszik leginkább megkülönböztethetővé az Allosaurustól.[42] Egy theropoda (Ceratosaurus vagy Torvosaurus) fognyomát viselő szeméremcsont lelet alapján az Allosaurus más húsevők zsákmányává is válhatott. A felsőtest alatt elhelyezkedő, és a lábak által védett csont, ami a csontváz egyik legerősebb része, jelzi hogy a példányból dögevők táplálkoztak.[77]
Ősbiológia[szerkesztés]
Élettörténet[szerkesztés]
A különböző korú Allosaurus fosszíliák bősége lehetővé teszi a tudósok számára az állat növekedésének vizsgálatát, és lehetséges élettartamának megbecslését. A maradványok között előfordulnak korai életszakaszból származók, akár tojások is – például Coloradoból olyan összetört tojások kerültek elő, amiket az Allosaurusénak tulajdonítanak.[2] A végtagok csontjainak szövettani elemzése alapján az Allosaurus élettartamának felső határa körülbelül 22–28 év lehetett, ami nagyjából megegyezik a többi nagy méretű theropoda, például a Tyrannosaurus maximális életkorával. Ezen elemzés alapján az is megállapítható, hogy az állatok körülbelül 15 évesen érték el a maximális méretüket, a testtömegüket évente mintegy 150 kilogrammal gyarapítva.[47]
A más dinoszauruszoknál, például a Tyrannosaurusnál és a Tenontosaurusnál fellelhető medulláris csontszövet legalább egy, a Cleveland-Lloyd lelőhelyről előkerült Allosaurus példány sípcsontjában is megtalálható. Napjainkban ez a csontszövet csak a nőstény madaraknál fordul elő tojásköltés idején, ugyanis a tojáshéjakhoz szükséges kalcium termelésére szolgál. Jelenléte az Allosaurusnál utalhat a példány nemére és ivarérettségére. A növekedési vonalak alapján ez az állat 10 éves lehetett a pusztulása idején, ami azt jelzi, hogy már a nemi érettség állapotában volt, mielőtt elérte volna a maximális testméretét.[78]
A csaknem teljes hátsó lábbal rendelkező kifejletlen példány felfedezése arra is bizonyítékul szolgál, hogy ezeknél az egyedeknél a lábak hossza aránylag nagyobb volt, és a láb alsó része (a lábszár és a talp) hosszabb volt a combnál. Ez a különbség arra utal, hogy a fiatalabb állatok gyorsabbak voltak, így más stratégiával vadásztak, mint a felnőttek; talán eleinte kisebb célpontokat szemeltek ki és vettek üldözőbe, majd felnőttként áttértek a nagyobb állatok lesből történő elejtésére.[48] A combcsont idővel vastagabbá és szélesebbé vált, keresztmetszete eltért a korábbi kör alaktól, így az izomtapadási pontjai is eltolódtak, az izmok rövidebbé váltak és a láb növekedése lelassult. A változások révén a láb képessé vált arra, hogy elbírja a felnőtt állat tömegét, melyet főként előrefelé kellett mozgatnia.[79]
Táplálkozás[szerkesztés]

Az őslénykutatók az Allosaurust nagyméretű állatokra vadászó aktív ragadozónak tekintik. A sauropodák maradványain talált sérülések, melyekbe beleillenek az Allosaurus fogai és a sauropoda fosszíliák közelében talált elhullajtott fogak, azt jelzik, hogy ezek az állatok a célpontjai lehettek, akár élő zsákmányra vadászott, akár dögevőként keresett táplálékot.[80] Egy Stegosaurus lelet drámai bizonyítékokkal szolgál egy támadására vonatkozóan, melyek között szerepel egy Allosaurus farokcsigolya egy részben begyógyult szúrás okozta sérüléssel, amibe beleillik a Stegosaurus faroktüskéje, valamint a Stegosaurus sérült nyaki lemeze, melynek U alakú bemetszésébe beleilleszthető az Allosaurus pofarésze.[81] Gregory S. Paul 1988-ban kijelentette, hogy az Allosaurus egymagában valószínűleg nem vadászott kifejlett sauropodákra, legfeljebb csapatban volt képes ilyesmire a mérsékelt méretű koponyája, az aránylag kis fogai és a tömege miatt, ami messze elmaradt a kortárs sauropodákétól.[9] Az is lehetséges, hogy a felnőttek helyett csupán a fiatal egyedekre vadászott.[6][61] Az 1990-es és 2000-es évek kutatásai talán egyéb magyarázattal is szolgálhatnak erre a kérdésre. Robert T. Bakker összehasonlította az Allosaurust a kainozoikum kardfogú húsevő emlőseivel, és olyan hasonlóságokat talált közöttük, mint például az állkapocs izomzat csökkenése és a nyakizmok megerősödése, valamint a szokatlanul nagyra nyitható állcsontok. Az Allosaurusnak azonban nem voltak kardfogai, így Bakker szerint más támadási módot kellett alkalmaznia, hogy kihasználhassa a feje és nyaka adta lehetőségeket, például a felső állcsont mentén levő rövid fogak fűrészként hatolhattak bele az áldozat húsába. Ez a fajta állcsont lehetővé teszi hogy az állat lecsapjon egy nagyobb méretű zsákmányra azzal a céllal, hogy meggyengítse az áldozatot.[50]

Más, az Allosaurus koponyáját érintő kutatások a végeselem analízis használatával hasonló eredményekre jutottak. A biomechanikai elemzés szerint a koponya nagyon erős volt, de aránylag kis harapáserővel rendelkezett. Csupán az állkapocs izmok segítségével 805–8724 N erőt lehetett képes kifejteni,[49][82] de a teljes koponya használatával már közel 55 500 newtont is elérhetett.[49] Azok a szerzők, akik szerint az Allosaurus a koponyáját szekerceként használva, nyitott szájjal csapott le a prédájára, kihasítva és kitépve belőle a húst, úgy vélik, hogy nem zúzta össze a csontokat, ellentétben a Tyrannosaurusszal, amelyről azt feltételezik, hogy erre is képes volt. Emellett azt is valószínűsítik, hogy a koponya felépítése lehetővé tehette, hogy az egyes zsákmányállatokat egyedi stratégiával támadta meg; kis tömege miatt alkalmas volt a gyors ornithopodák elejtésére, de elég erős volt ahhoz is, hogy lecsapjon vele az olyan nagyobb méretű állatokra, mint a stegosauridák vagy a sauropodák.[49] Egyes kutatók azonban vitatják ezt az elképzelést, arra hivatkozva, hogy nem találtak modern példát a szekerceként való koponyahasználatra, bár úgy vélik, hogy a testrész nyitott állapotában is elég erős volt ahhoz, hogy a vergődő préda által okozott igénybevételt elviselje.[83] A többféle támadási stratégiát lehetővé tevő koponya mellett érvelő szerzők szerint, bár az Allosaurusnak nincs mai megfelelője, a fogsor jó illeszkedése lehetővé tette a szekerceként való lecsapást, emellett pedig az elrendezése segítette a szájpadlás védelmét és csökkentette a rá ható erőt.[84] Egyéb lehetőségként az is felmerült, hogy a nagyobb állatokról az Allosaurushoz hasonló theropodák „legelték” a húst, azaz kiharaptak egy-egy darabot az élő állatokból, ami elegendő volt a fennmaradásukhoz, így nem volt szükségük arra, hogy megöljék a prédát. Ez a stratégia azt is lehetővé tenné, hogy a sérülésből felgyógyult áldozat a későbbiekben újra élelemforrásként szolgáljon.[13] Egy állítás szerint az ornithopodák jóval könnyebben elérhető zsákmányállatok voltak, és az Allosaurusok talán a modern nagymacskákhoz hasonlóan végeztek velük; mellső lábaikkal megragadták, majd a légcső összezúzása érdekében többször torkon harapták az állatot.[6] Ez összhangban áll az erős és a zsákmány megtartására képes mellső lábakra utaló bizonyítékokkal.[20]
A táplálkozás szempontjából fontosak a szemek, valamint a mellső és hátsó végtagok is. Az Allosaurus koponyájának formája és szemeinek elhelyezkedése 20°-os szögre korlátozza a binokuláris látást, ami valamivel kisebb annál, ami a ma élő krokodiloknál mérhető. Ez elég lehetett ahhoz, hogy az Allosaurus a krokodilokhoz hasonlóan képes legyen megítélni a zsákmány távolságát és a támadás megfelelő pillanatát.[85] Emellett pedig arra utal, hogy az állat a krokodilokhoz hasonlóan lesből támadt.[86] A mellső lábak a többi theropodáéhoz hasonlítva, a zsákmány adott távolságból történő megragadására, illetve fogva tartására alkalmasak,[20] a kampószerű karmok mindkét esetben segítséget nyújthattak.[19] Az Allosaurus csúcssebességét 30–55 km/h-ra becsülik.[87]
Szociális viselkedés[szerkesztés]
Az Allosaurust a félig tudományos és szórakoztató irodalomban sokáig olyan állatként ismertették, amely csapatosan vadászott a sauropodákra és más nagy méretű dinoszauruszokra.[11][28][61] Robert T. Bakker kiterjesztette a szociális viselkedést a szülői gondoskodásra vonatkozó elképzelésekkel, ugyanis az elhullajtott Allosaurus fogak és a nagy testű zsákmányállatok megrágott csontjai alapján úgy vélte, hogy a felnőtt egyedek élelmet hordtak a búvóhelyükre, hogy a kicsinyeiket táplálják, illetve hogy elrejtsék a húst a dögevők elől.[52] Habár kevés bizonyíték létezik a theropodák csapatban való tevékenykedésére,[13] az azonos fajba tartozó egyedek közötti szociális kölcsönhatás részben ellenséges lehetett, ami a hasi bordákon[15] és a koponyákon levő sérülésekből látható (erre szolgálhat bizonyítékul a Labrosaurus ferox kórosan elváltozott állkapcsa is). Az efféle fejharapások talán a dominancia eldöntésének módszerei voltak a csapatban vadászó állatok között, vagy a területi viták rendezésében játszottak szerepet.[88]
Az Allosaurus talán csapatban vadászott,[89] de egy újkeletű kutatás szerint az Allosaurusra és más theropodákra inkább a meglehetősen agresszív viselkedés volt jellemző, mint a fajtársak közti együttműködés. A kérdéses tanulmány a csapatban való vadászattal kapcsolatban megemlíti, hogy a vadászat e formája, ami lehetővé tenné a ragadozó egyedeknél nagyobb méretű zsákmány elejtését, a theropoda dinoszauruszoknál nagyon ritka lehetett, ahogy az a gerincesekre és köztük a modern diapsida húsevőkre (a gyíkokra, a krokodilokra és a madarakra) is jellemző, melyek jellemzően territoriálisak, és megölik vagy akár kannibálként el is fogyasztják a betolakodó fajtársakat, illetve azokat a kisebb példányokat, amelyek egy esetleges közös zsákmányból előttük próbálnak enni. Ezen értelmezés szerint például a Cleveland-Lloyd lelőhelyen talált Allosaurus egyedek nem a csapatos vadászat miatt gyűltek össze, hanem azért, hogy egy másik sérült vagy elpusztult példányból táplálkozzanak, de némelyiket megölték eközben. Ez megmagyarázná a fiatal és kifejletlen egyedek magas elhullási arányát, ami hasonló ahhoz, ami napjainkban a krokodilok és komodói varánuszok táplálkozási helyein tapasztalható. Ez a fajta értelmezés Bakker búvóhelyekkel kapcsolatos elméleténél is használható.[90] Az Allosaurusok kannibalizmusára létezik bizonyíték, például találtak elhullajtott fogakat az összetört bordák között, és feltételezett fognyomokat egy lapockacsonton,[91] valamint csontok között heverő, elfogyasztott Allosaurus csontvázakat Bakker búvóhely lelőhelyein.[76]
Agy és érzékszervek[szerkesztés]
Az Allosaurus agya egy koponyaöntvény spirális CT letapogatása alapján jobban hasonlít a krokodilok agyához, mint más ma élő archosaurusokéhoz, például a madarakéhoz. Az előcsarnoki rendszer szerkezete azt jelzi, hogy a koponya nagyjából vízszintesen helyezkedett el, nem erősen előre- vagy hátrabillentett helyzetben. A belső fül szerkezete szintén a krokodiléra emlékeztet, és ebből következően az állat valószínűleg a mély frekvenciatartományban hallott a legjobban, a finom hangokat pedig nehezen kezelte. A nagy méretű szaglógumó jó szaglást biztosított, de a szagfelismerést végző terület aránylag kicsi volt.[51]
Popkulturális hatás[szerkesztés]

A popkultúrában a Tyrannosaurus mellett az Allosaurus jelképezi az ideális nagy testű húsevő dinoszauruszt. Gyakran fordul elő a múzeumokban, főként a Cleveland-Lloyd dinoszaurusz lelőhelyen végzett ásatások óta; 1976-tól kezdődően három kontinens nyolc országának 38 múzeuma jutott innen Allosaurus maradványokhoz vagy róluk készült öntvényekhez.[4]
1988-ban az egyesült államokbeli Utahban az Allosaurust az állam fosszíliájának nyilvánították.[92]
Az Allosaurust a 20. század eleje óta sokszor ábrázolták a populáris kultúrában:
- Elsőként 1912-ben, Arthur Conan Doyle Az elveszett világ című regényében és a belőle 1925-ben készült némafilm adaptációban jelent meg, amely az első egész estés produkció, amiben dinoszauruszok láthatók (egyebek mellett a Tyrannosaurus is felbukkan).[93]
- Főszerepet kapott az 1956-os The Beast of Hollow Mountain (Az üreges hegy szörnye) című filmben.[94]
- Az 1969-es A tiltott völgy (The Valley of Gwangi) című filmben szereplő állatot Ray Harryhausen ugyan a Tyranosaurusról mintázta, az mégis jobban hasonlít egy Allosaurusra (egy DVD interjúban elhangzottak alapján néha összetévesztette őket: „Mindkettő húsevő, mindkettő zsarnok… az egyik egy kicsit nagyobb, mint a másik.”).
- Emellett az Allosaurus megjelent az 1966-os Egymillió évvel időszámításunk előtt (One Million Years B.C.) című filmben,
- Edgar Rice Burroughs The Land That Time Forgot (A föld, melyet elfeledett az idő) című regényének 1975-ös filmadaptációjában.
- A BBC által készített dokumentumfilm-sorozat, a Dinoszauruszok, a Föld urai Nagy Al balladája című különkiadása mellett a sorozat második részében több, Diplodocusra vadászó példány is feltűnik, az ötödik részben pedig egy Leaellynasaurát elejtő kis méretű antarktiszi faj látható. E sorozat folytatásának (Szörnyek bolygója – Walking with Monsters) végén az Allosaurus megjelenése jelzi a dinoszauruszok uralmának kezdetét.
- A Discovery Channel Dinoszauruszok, az ősvilág urai (When Dinosaurs Roamed America, 2001) című dokumentumfilmje is bemutatja ezt az állatot.
- A History csatorna 2008-ban készült Őshüllők arénája (Jurassic Fight Club) dokumentumfilm sorozatban is felbukkan a Ceratosaurus ellenfeleként.
- A Dinosaucers című rajzfilmsorozat „Allo” nevű főhőse egy Allosaurus.
- Ez az állat több képregényben is szerepel, például az Age of Reptiles és a Kázmér és Huba című sorozatokban, de feltűnik az Irregular Webcomic! és a Questionable content című internetes képregényekben is.
- Ez a dinoszaurusz több számítógépes játékba is bekerült, találkozhatunk vele a Jurassic Park: Operation Genesis és a Dino Crisis 2 állatai között.
Jegyzetek[szerkesztés]
- ↑ a b Liddell & Scott. Greek–English Lexicon, Abridged Edition, Oxford: Oxford University Press (1980). ISBN 0-19-910207-4. OCLC 17396377 187441362 213803616 217157617 225305941 237142939 43379590 78588771
- ↑ a b c d Glut, Donald F.. Allosaurus, Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co, 105–117. o. (1997). ISBN 0-89950-917-7
- ↑ a b c d e Mortimer, Mickey: And the Largest Theropod Is…. Dinosaur Mailing List. The Dinosaur Mailing List, 2003. július 21. [2010. március 25-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
- ↑ a b c d e f g h i j k l m n o p q r s Madsen, James H., Jr.. Allosaurus fragilis: A Revised Osteology, 2nd ed., Utah Geological Survey Bulletin 109, Salt Lake City: Utah Geological Survey [1976] (1993)
- ↑ a b Foster, John R.. Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A., New Mexico Museum of Natural History and Science Bulletin 23. Albuquerque: New Mexico Museum of Natural History and Science, 37. o. (2003)
- ↑ a b c d Foster, John. Allosaurus fragilis, Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Bloomington, Indiana: Indiana University Press, 170-176. o. (2007). ISBN 978-0-253-34870-8. OCLC 77830875
- ↑ a b Chure, Daniel J.. A new species of Allosaurus from the Morrison Formation of Dinosaur National Monument (Utah–Colorado) and a revision of the theropod family Allosauridae, Ph.D. dissertation. Columbia University (2000)
- ↑ Foster, John. Jurassic West: the Dinosaurs of the Morrison Formation and Their World. Bloomington, Indiana: Indiana University Press, 117. o. (2007)
- ↑ a b c d e f g h i j k l m n Paul, Gregory S.. Genus Allosaurus, Predatory Dinosaurs of the World. New York: Simon & Schuster, 307–313. o. (1988). ISBN 0-671-61946-2
- ↑ Molnar, Ralph E. (1977). „Analogies in the evolution of combat and display structures in ornithopods and ungulates”. Evolutionary Theory 3, 165–190. o.
- ↑ a b c Norman, David B.. 'Carnosaurs', The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. New York: Crescent Books, 62–67. o. (1985). ISBN 0-517-468905
- ↑ Paul, Gregory S.. Predatory Dinosaurs of the World, 91-93. o. (1988).
- ↑ a b c d e f g h i j k l m n Holtz, Thomas R., Jr., Molnar, Ralph E.; and Currie, Philip J..szerk.: Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka: The Dinosauria, 2nd ed., Berkeley: University of California Press, 71–110. o. (2004). ISBN 0-520-24209-2
- ↑ Paul, Gregory S.. Predatory Dinosaurs of the World, 277. o. (1988)
- ↑ a b c d Chure, Daniel J. (2000). „Observations on the morphology and pathology of the gastral basket of Allosaurus, based on a new specimen from Dinosaur National Monument”. Oryctos 3, 39–37. o. ISSN 1290-4805.
- ↑ Chure, Daniel J., and Madsen, James H. (1996). „On the presence of furculae in some non-maniraptoran theropods”. Journal of Vertebrate Paleontology 16 (3), 573–577. o.
- ↑ Middleton, Kevin M., and Gatesy, Stephen M. (2000). „Theropod forelimb design and evolution” (PDF). Zoological Journal of the Linnean Society 128, 149–187. o. DOI:10.1006/zjls.1998.0193. (Hozzáférés: 2009. március 26.)
- ↑ Utah State Fossil: Theropod Dinosaur (Allosaurus fragilis). [2009. április 1-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
- ↑ a b c d e Gilmore, Charles W. (1920). „Osteology of the carnivorous dinosauria in the United States National Museum, with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus”. Bulletin of the United States National Museum 110, 1–159. o.
- ↑ a b c Carpenter, Kenneth (2002). „Forelimb biomechanics of nonavian theropod dinosaurs in predation” (PDF). Senckebergiana lethaea 82 (1), 59–76. o. [2007. szeptember 27-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
- ↑ Paul, Gregory S.. Predatory Dinosaurs of the World, 113, 310-311. o. (1988)
- ↑ a b Marsh, Othniel Charles (1878). „Notice of new dinosaurian reptiles”. American Journal of Science and Arts 15, 241–244. o.
- ↑ von Huene, Friedrich (1926). „The carnivorous Saurischia in the Jura and Cretaceous formations, principally in Europe”. Revista del Museo de La Plata 29, 35–167. o.
- ↑ Romer, Alfred S.. Osteology of the Reptiles. Chicago: University of Chicago Press (1956). ISBN 0-89464985-X
- ↑ Romer, Alfred S.. Vertebrate Paleontology, Third Edition, Chicago: University of Chicago Press (1966). ISBN 0-7167-1822-7
- ↑ Steel, R. (1970). „Part 14. Saurischia. Handbuch der Paläoherpetologie/Encyclopedia of Paleoherpetology”. Gustav Fischer Verlag, Stuttgart, 1–87. o.
- ↑ Walker, Alick D. (1964). „Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs”. Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences 248 (744), 53–134. o. DOI:10.1098/rstb.1964.0009. (Hozzáférés: 2009. március 26.)
- ↑ a b Lambert, David, and the Diagram Group. Allosaurids, A Field Guide to Dinosaurs. New York: Avon Books, 80–81. o. (1983). ISBN 0-380-83519-3
- ↑ Lambert, David, and the Diagram Group. Allosaurids, The Dinosaur Data Book. New York: Avon Books, 130. o. (1990). ISBN 0-380-75896-3
- ↑ Paul, Gregory S.. The allosaur-tyrannosaur group, Predatory Dinosaurs of the World, 301–347. o. (1988)
- ↑ Holtz, Thomas R., Jr. (1994). „The phylogenetic position of the Tyrannosauridae: Implications for theropod systematics”. Journal of Paleontology 68 (5), 1100–1117. o. (Hozzáférés: 2009. március 26.)
- ↑ Leidy, Joseph (1870). „Remarks on Poicilopleuron valens, Clidastes intermedius, Leiodon proriger, Baptemys wyomingensis, and Emys stevensonianus”. Proceedings of the Academy of Natural Sciences, Philadelphia 22, 3–4. o.
- ↑ Leidy, Joseph (1873). „Contribution to the extinct vertebrate fauna of the western territories”. Report of the U.S. Geological Survey of the Territories I, 14–358. o.
- ↑ a b Marsh, Othniel Charles (1877). „Notice of new dinosaurian reptiles from the Jurassic formation”. American Journal of Science and Arts 14, 514–516. o.
- ↑ Creisler, Ben: Dinosauria Translation and Pronunciation Guide A. Dinosauria On-Line, 2003. július 7. [2010. január 17-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
- ↑ Marsh, Othniel Charles (1879). „Principal characters of American Jurassic dinosaurs. Part II”. American Journal of Science, Series 3 17, 86–92. o.
- ↑ Cope, Edward Drinker (1878). „A new opisthocoelous dinosaur”. American Naturalist 12 (6), 406. o. DOI:10.1086/272127.
- ↑ Norell, Mark A., Gaffney, Eric S.; and Dingus, Lowell. Discovering Dinosaurs in the American Museum of Natural History. New York: Knopf, 112–113. o. (1995). ISBN 0-679-43386-4
- ↑ Breithaupt, Brent H., Chure, Daniel J.; and Southwell, Elizabeth H. (1999). „AMNH 5753: The world's first free-standing theropod skeleton”. Journal of Vertebrate Paleontology 19 (3, Suppl.), 33A. o.
- ↑ Williston, Samuel Wendell (1878). „American Jurassic dinosaurs”. Transactions of the Kansas Academy of Science 6, 42–46. o.
- ↑ Williston, Samuel Wendell (1901). „The dinosaurian genus Creosaurus, Marsh”. American Journal of Science, series 4 11 (11), 111–114. o.
- ↑ a b Henderson, Donald M. (1998). „Skull and tooth morphology as indicators of niche partitioning in sympatric Morrison Formation theropods” (PDF). Gaia 15, 219–266. o. [2011. július 19-i dátummal az eredetiből archiválva]. ISSN 0871-5424. (Hozzáférés: 2009. március 26.)
- ↑ Stokes, William J. (1945). „A new quarry for Jurassic dinosaurs”. Science 101 (2614), 115–117. o. DOI:10.1126/science.101.2614.115-a. PMID 17799203.
- ↑ Hunt, Adrian P, Lucas, Spencer G.; Krainer, Karl; and Spielmann, Justin.szerk.: Foster, John R.; and Lucas, Spencer G.: The taphonomy of the Cleveland-Lloyd Dinosaur Quarry, Upper Jurassic Morrison Formation, Utah: a re-evaluation, Paleontology and Geology of the Upper Jurassic Morrison Formation, New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 57–65. o. (2006)
- ↑ Loewen, Mark A., Sampson, Scott D.; Carrano, Matthew T.; and Chure, Daniel J. (2003). „Morphology, taxonomy, and stratigraphy of Allosaurus from the Upper Jurassic Morrison Formation”. Journal of Vertebrate Paleontology 23 (3, Suppl.), 72A. o.
- ↑ a b Smith, David K. (1998). „A morphometric analysis of Allosaurus”. Journal of Vertebrate Paleontology 18 (1), 126–142. o.
- ↑ a b Bybee, Paul J., Lee, A.H.; and Lamm, E.-T. (2006). „Sizing the Jurassic theropod dinosaur Allosaurus: Assessing growth strategy and evolution of ontogenetic scaling of limbs”. Journal of Morphology 267 (3), 347–359. o. DOI:10.1002/jmor.10406.
- ↑ a b Foster, John R., and Chure, Daniel J..szerk.: Foster, John R.; and Lucas, Spencer G.: Hindlimb allometry in the Late Jurassic theropod dinosaur Allosaurus, with comments on its abundance and distribution, Paleontology and Geology of the Upper Jurassic Morrison Formation, New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 119–122. o. (2006)
- ↑ a b c d Rayfield, Emily J., Norman, David B.; Horner, Celeste C.; Horner, John R.; Smith, Paula May; Thomason, Jeffrey J.; and Upchurch, Paul (2001). „Cranial design and function in a large theropod dinosaur”. Nature 409, 1033–1037. o. DOI:10.1038/35059070.
- ↑ a b Bakker, Robert T. (1998). „Brontosaur killers: Late Jurassic allosaurids as sabre-tooth cat analogues” (PDF). Gaia 15, 145–158. o. [2007. október 31-i dátummal az eredetiből archiválva]. ISSN 0871-5424. (Hozzáférés: 2009. március 26.)
- ↑ a b Rogers, Scott W. (1999). „Allosaurus, crocodiles, and birds: Evolutionary clues from spiral computed tomography of an endocast” (PDF). The Anatomical Record 257 (5), 163–173. o. [2020. április 1-i dátummal az eredetiből archiválva]. DOI:<162::AID-AR5>3.0.CO;2-W 10.1002/(SICI)1097-0185(19991015)257:5<162::AID-AR5>3.0.CO;2-W. (Hozzáférés: 2009. március 26.)
- ↑ a b Bakker, Robert T..szerk.: Wolberg, Donald L.; Sump, Edmund; and Rosenberg, Gary D.: Raptor Family values: Allosaur parents brought giant carcasses into their lair to feed their young, Dinofest International, Proceedings of a Symposium Held at Arizona State University. Philadelphia: Academy of Natural Sciences, 51–63. o. (1997). ISBN 0-935868-94-1
- ↑ Chure, Daniel J..szerk.: Ailing Sun and Yuangqing Wang: A reassessment of the gigantic theropod Saurophagus maximus from the Morrison Formation (Upper Jurassic) of Oklahoma, USA, Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers. Beijing: China Ocean Press, 103–106. o. (1995). ISBN 7-5027-3898-3
- ↑ Pérez-Moreno, B.P., Chure, D.J.; Pires, C.; Silva, C.M.; Santos, V.; Dantas, P.; Póvoas, L.; Cachăo, M.; Sanz, J.L.; and Galopim De Carvalho, A.M. (1999). „On the presence of Allosaurus fragilis (Theropoda: Carnosauria) in the Upper Jurassic of Portugal: First evidence of an intercontinental dinosaur species” (PDF). Journal of the Geological Society 156 (3), 449–452. o. [2021. június 6-i dátummal az eredetiből archiválva]. DOI:10.1144/gsjgs.156.3.0449. (Hozzáférés: 2009. március 26.)
- ↑ a b Breithaupt, Brent.szerk.: Brown, C.E.; Kirkwood, S.C.; and Miller, T.S.: The discovery of a nearly complete Allosaurus from the Jurassic Morrison Formation, eastern Bighorn Basin, Wyoming, Forty-Seventh Annual Field Conference Guidebook. Casper, Wyoming: Wyoming Geological Association, 309–313. o. (1996). OCLC 36004754
- ↑ a b Howe Dinosaur Quarry – Wyoming’s Jurassic Treasure. GeoScience Adventures, 2007. július 24. (Hozzáférés: 2009. március 26.)
- ↑ Piotrowska, Anna (2001. december 9.). „Tropami dinozaurów” (lengyel nyelven). Newsweek (Polish edition), 68–70. o.
- ↑ a b Breithaupt, Brent H.: The case of "Big Al" the Allosaurus: a study in paleodetective partnerships. [2010. január 17-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
- ↑ a b Hanna, Rebecca R. (2002). „Multiple injury and infection in a sub-adult theropod dinosaur (Allosaurus fragilis) with comparisons to allosaur pathology in the Cleveland-Lloyd dinosaur quarry collection”. Journal of Vertebrate Paleontology 22 (1), 76–90. o. DOI:[0076:MIAIIA2.0.CO;2 10.1671/0272-4634(2002)022[0076:MIAIIA]2.0.CO;2].
- ↑ a b Mateus, Octávio, Walen, Aart; and Antunes, Miguel Telles.szerk.: Foster, John R.; and Lucas, Spencer G.: The large theropod fauna of the Lourinha Formation (Portugal) and its similarity to that of the Morrison Formation, with a description of a new species of Allosaurus, Paleontology and Geology of the Upper Jurassic Morrison Formation, New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 123–129. o. (2006)
- ↑ a b c Lessem, Don, and Glut, Donald F.. Allosaurus, The Dinosaur Society's Dinosaur Encyclopedia. Random House, 19–20. o. (1993). ISBN 0-679-41770-2. OCLC 30361459
- ↑ a b Smith, David K. (1996). „A discriminant analysis of Allosaurus population using quarries as the operational units”. Museum of Northern Arizona Bulletin 60, 69–72. o.
- ↑ Smith, David K. (1999). „Patterns of size-related variation within Allosaurus”. Journal of Vertebrate Paleontology 19 (2), 402–403. o.
- ↑ Malafaia, Elisabete, Dantas, Pedro; Ortega, Francisco; and Escaso, Fernando (2007). „Nuevos restos de Allosaurus fragilis (Theropoda: Carnosauria) del yacimiento de Andrés (Jurásico Superior; centro-oeste de Portugal)” (spanyol with english abstract nyelven) (PDF). Cantera Paleontológica, 255–271. o. (Hozzáférés: 2009. március 26.)
- ↑ Janensch, Werner (1925). „Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas” (német nyelven). Palaeontographica, Supplement 7 1, 1–99. o.
- ↑ Rauhut, Oliver W.M. (2005). „Post-cranial remains of 'coelurosaurs' (Dinosauria, Theropoda) from the Late Jurassic of Tanzania”. Geological Magazine 142 (1), 97–107. o. DOI:10.1017/S0016756804000330.
- ↑ Marsh, Othniel Charles (1884). „Principal characters of American Jurassic dinosaurs. Part VIII”. American Journal of Science, Series 3 27, 329–340. o.
- ↑ Madsen, James H., and Welles, Samuel P.. Ceratosaurus (Dinosauria, Theropoda), a Revised Osteology, Miscellaneous Publication, 00-2. Utah Geological Survey (2000)
- ↑ Molnar, Ralph E., Flannery, Timothy F.; and Rich, Thomas H.V. (1981). „An allosaurid theropod dinosaur from the Early Cretaceous of Victoria, Australia”. Alcheringa 5, 141–146. o. DOI:10.1080/03115518108565427.
- ↑ Riabinin, Anatoly Nikolaenvich (1914). „Zamtka o dinozavry ise Zabaykalya” (orosz nyelven). Trudy Geologichyeskago Muszeyah Imeni Petra Velikago Imperatorskoy Academiy Nauk 8 (5), 133–140. o.
- ↑ Foster, John R.. Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A., New Mexico Museum of Natural History and Science Bulletin, 23. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 29. o. (2003)
- ↑ Russell, Dale A.. An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press, 175–176. o. (1989). ISBN 1-55971-038-1
- ↑ Carpenter, Kenneth.szerk.: Foster, John R.; and Lucas, Spencer G.: Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus, Paleontology and Geology of the Upper Jurassic Morrison Formation [archivált változat] (PDF), New Mexico Museum of Natural History and Science Bulletin, 36, Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 131–138. o. (2006). Hozzáférés ideje: 2009. március 26. [archiválás ideje: 2007. december 2.] Archivált másolat. [2007. december 2-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. április 1.)
- ↑ Chure, Daniel J., Litwin, Ron; Hasiotis, Stephen T.; Evanoff, Emmett; and Carpenter, Kenneth.szerk.: Foster, John R.; and Lucas, Spencer G.: The fauna and flora of the Morrison Formation: 2006, Paleontology and Geology of the Upper Jurassic Morrison Formation, New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 233–248. o. (2006)
- ↑ a b Mateus, Octávio.szerk.: Foster, John R.; and Lucas, Spencer G.: Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhă and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison, Paleontology and Geology of the Upper Jurassic Morrison Formation, New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science, 223–231. o. (2006)
- ↑ a b Bakker, Robert T., and Bir, Gary.szerk.: Currie, Philip J.; Koppelhus, Eva B.; Shugar, Martin A.; and Wright, Joanna L.: Dinosaur crime scene investigations: theropod behavior at Como Bluff, Wyoming, and the evolution of birdness, Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Bloomington and Indianapolis: Indiana University Press, 301–342. o. (2004). ISBN 0-253-34373-9
- ↑ Chure, Daniel J., Fiorillo, Anthony R.; and Jacobsen, Aase (2000). „Prey bone utilization by predatory dinosaurs in the Late Jurassic of North America, with comments on prey bone use by dinosaurs throughout the Mesozoic” (PDF). Gaia 15, 227–232. o. [2011. július 19-i dátummal az eredetiből archiválva]. ISSN 0871-5424. (Hozzáférés: 2009. március 26.)
- ↑ Lee, Andrew H., and Werning, Sarah (2008). „Sexual maturity in growing dinosaurs does not fit reptilian growth models”. Proceedings of the National Academy of Sciences 105 (2), 582–587. o. DOI:10.1073/pnas.0708903105. PMID 18195356.
- ↑ Loewen, Mark A., Carrano, Matthew T.; and Sampson, Scott D. (2002). „Ontogenetic changes in hindlimb musculature and function in the Late Jurassic theropod Allosaurus”. Journal of Vertebrate Paleontology 22 (3, Suppl.), 80A. o.
- ↑ Fastovsky, David E.; and Smith, Joshua B.. Dinosaur Paleoecology, The Dinosauria, 2nd edition, 614–626. o. (2004)
- ↑ Kenneth, Carpenter, Sanders, Frank; McWhinney, Lorrie A.; and Wood, Lowell.szerk.: Carpenter, Kenneth (ed.): Evidence for predator-prey relationships: Examples for Allosaurus and Stegosaurus, The Carnivorous Dinosaurs [archivált változat] (PDF), Bloomington and Indianapolis: Indiana University Press, 325–350. o. (2005). ISBN 0-253-34539-1. Hozzáférés ideje: 2009. március 26. [archiválás ideje: 2009. március 26.]
- ↑ K.T. Bates, P.L. Falkingham (2012). „Estimating maximum bite performance in Tyrannosaurus rex using multi-body dynamics”. Biological Letters 8 (4), 660–664. o. [2018. augusztus 19-i dátummal az eredetiből archiválva]. DOI:10.1098/rsbl.2012.0056. PMID 22378742. (Hozzáférés: 2018. szeptember 14.)
- ↑ Frazzetta, T.H., and Kardong, Kenneth V. (2002). „Prey attack by a large theropod dinosaur”. Nature 416, 387–388. o. DOI:10.1038/416387a.
- ↑ Rayfield, Emily J., Norman, David B.; and Upchurch, Paul (2002). „Prey attack by a large theropod dinosaur: Response to Frazzetta and Kardong, 2002”. Nature 416, 388. o. DOI:10.1038/416388a.
- ↑ Stevens, Kent A. (2006). „Binocular vision in theropod dinosaurs”. Journal of Vertebrate Paleontology 26 (2), 321–330. o. DOI:[321:BVITD2.0.CO;2 10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2].
- ↑ „Evolve: Eyes”. History channel Evolve. [1]. Archiválás az eredetiről (2009. június 28.). Hozzáférés: 2009. március 26.
- ↑ Christiansen, Per (1998). „Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential” (PDF). Gaia 15, 241–255. o. [2008. október 31-i dátummal az eredetiből archiválva]. ISSN 0871-5424. (Hozzáférés: 2009. március 26.)
- ↑ Tanke, Darren H., and Currie, Philip J. (1998). „Head-biting behavior in theropod dinosaurs: Paleopathological evidence” (PDF). Gaia (15), 167–184. o. [2006. január 11-i dátummal az eredetiből archiválva]. ISSN 0871-5424. (Hozzáférés: 2009. március 26.)
- ↑ Currie, Philip J..szerk.: Farlow, James; and Brett-Surman, M.K.: Theropods, The Complete Dinosaur. Indiana: Indiana University Press, 228. o. (1999). ISBN 0253213134. Hozzáférés ideje: 2009. március 26.
- ↑ Roach, Brian T., and Brinkman, Daniel L. (2007). „A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History 48 (1), 103–138. o. DOI:[103:AROCPH2.0.CO;2 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2].
- ↑ Goodchild Drake, Brandon (2004). „A new specimen of Allosaurus from north-central Wyoming”. Journal of Vertebrate Paleontology 24 (3, Suppl.), 65A. o.
- ↑ Utah Symbols – State Fossil. State of Utah. [2016. március 3-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
- ↑ Glut, Donald F., Brett-Surman, Michael K..szerk.: Farlow, James; and Brett-Surman, Michael K.: Dinosaurs and the media, The Complete Dinosaur. Bloomington and Indianapolis: Indiana University Press, 675–706. o. (1997). ISBN 0-253-33349-0
- ↑ Lambert, David, and the Diagram Group. Allosaurids, The Dinosaur Data Book. New York: Avon Books, 299. o. (1990). ISBN 0-380-75896-3
Fordítás[szerkesztés]
- Ez a szócikk részben vagy egészben az Allosaurus című angol Wikipédia-szócikk ezen változatának fordításán alapul. Az eredeti cikk szerkesztőit annak laptörténete sorolja fel. Ez a jelzés csupán a megfogalmazás eredetét és a szerzői jogokat jelzi, nem szolgál a cikkben szereplő információk forrásmegjelöléseként.
További információk[szerkesztés]
Magyar nyelven[szerkesztés]
- Atlanti-óceánt átszelő dinoszauruszok?. [2006. május 29-i dátummal az eredetiből archiválva]. (Hozzáférés: 2021. augusztus 18.)
- Allosaurus (magyar nyelven). hanghatásokkal színesített oldal. setaadinoszauruszokkal.hu. [2010. október 5-i dátummal az eredetiből archiválva]. (Hozzáférés: 2011. május 19.)
Angol nyelven[szerkesztés]
- Allosaurus, the story of "Big Al". [2007. február 14-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
- Specimens, discussion, and references pertaining to Allosaurus. [2012. december 20-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
- Allosaurus. (Hozzáférés: 2009. március 26.)
- Public Pioneer, Utah State Fossil, Allosaurus. [2016. március 3-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
- Restoration of MOR 693 ("Big Al"). [2007. szeptember 26-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
- muscle and organ restoration. [2007. október 7-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
- Allosaurus species. Dinosaur Mailing List, 1995. november 30. [2005. március 24-i dátummal az eredetiből archiválva]. (Hozzáférés: 2009. március 26.)
{kind=link}
Kapcsolódó szócikkek[szerkesztés]