
Albertosaurus
| Evolúciós időszak: késő kréta 72–70 Ma | ||||||||||||||||||||||||
 Az Albertosaurus rekonstrukciója az albertai Royal Tyrrell Museumban
| ||||||||||||||||||||||||
| Természetvédelmi státusz | ||||||||||||||||||||||||
| Fosszilis | ||||||||||||||||||||||||
| Rendszertani besorolás | ||||||||||||||||||||||||
| ||||||||||||||||||||||||
| Szinonimák | ||||||||||||||||||||||||
| ||||||||||||||||||||||||
| Fajok | ||||||||||||||||||||||||
| ||||||||||||||||||||||||
| Hivatkozások | ||||||||||||||||||||||||
A Wikifajok tartalmaz Albertosaurus témájú rendszertani információt. A Wikimédia Commons tartalmaz Albertosaurus témájú médiaállományokat és Albertosaurus témájú kategóriát. |
Az Albertosaurus (nevének jelentése 'Alberta gyík') a tyrannosaurida Theropoda dinoszauruszok egyik neme, amely Észak-Amerika nyugati részén élt a késő kréta időszakban, több mint 70 millió évvel ezelőtt. Típusfaja, az A. sarcophagus élettere a kanadai Alberta tartományra korlátozódott, melyről a nemet elnevezték. Az őslénykutatók nem értenek egyet abban, hogy hány faj tartozik ide, egyesek szerint a Gorgosaurus libratus a nem második faja.
Az Albertosaurus más tyrannosauridákhoz hasonlóan két lábon járó ragadozó volt, amely aránylag apró, két ujjú mellső lábakkal és súlyos fejjel rendelkezett, amiben több tucatnyi nagy méretű, éles fog helyezkedett el. A helyi ökoszisztémán belül a tápláléklánc csúcsán állhatott. Habár aránylag nagy méretű volt egy theropodához képest, mégis sokkal kisebb volt ismertebb rokonánál a Tyrannosaurusnál, a tömege legfeljebb 2,5 tonna lehetett.
Több mint harminc egyed fosszíliáját fedezték fel, ami lehetővé tette a kutatók számára, hogy a legtöbb theropodáénál részletesebb képet kapjanak az Albertosaurus anatómiájáról. Az egyik lelőhelyen talált 22 példány bizonyítékkal szolgál arra vonatkozóan, hogy csapatban éltek, és lehetővé teszi egyedfejlődési és populációbiológiai szempontból történő tanulmányozásukat, ami a kevésbé ismert dinoszauruszok esetében lehetetlen.
Anatómia[szerkesztés]
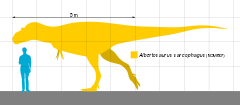

Az Albertosaurus kisebb volt az olyan óriási méretű tyrannosauridáknál, mint a Tarbosaurus és a Tyrannosaurus. Az átlagos kifejlett egyedek hossza 8–9 méter volt,[1][2][3] de egyes ritka, idős példányok meghaladták a 10 métert is.[4] Számos egymástól független, különböző módszerrel készült tömegbecslés létezik, melyek alapján egy felnőtt Albertosaurus tömege körülbelül 1,3–2,5 tonna lehetett.[3][5][6][7][8]
Az Albertosaurus súlyos koponyája egy rövid, S alakú nyakon helyezkedett el, melynek hossza a nagyobb felnőttek esetében 1 méter körül volt.[9] A koponya tömegét széles nyílások (fenestrae-k) csökkentették, amik egyúttal izomtapadási pontként is szolgáltak, és helyet biztosítottak az érzékszervek (szemek, orrüreg, hallószervek) számára. Hosszú állkapcsaiban több mint 60 banán alakú fog helyezkedett el; a nagyobb tyrannosauridáknak ennél kevesebb volt.
A legtöbb theropodától eltérően az Albertosaurus és más tyrannosauridák különböző helyen levő fogainál alaki eltérés (heterodontia) figyelhető meg. A premaxilláris fogak a felső állcsont elején sokkal kisebbek voltak a többinél, egymáshoz közelebb helyezkedtek el, és a keresztmetszetük D alakú volt.[2] A szemek felett rövid, csontos kinövések voltak, amik talán élénk színűek lehettek, és a párkeresés során a figyelem felkeltésére szolgáltak.[10]
A tyrannosauridák külseje hasonló volt, és az Albertosaurus sem kivétel ez alól. A theropodákra jellemzően két lábon járt, felsőtestét és nehéz fejét pedig hosszú farka tartotta egyensúlyban. A tyrannosauridák mellső lába azonban rendkívül kicsi volt a testméretükhöz képest, és csak két ujj volt rajta. A hátsó lábak hosszúak voltak, és egy-egy négyujjú lábfejben végződtek. Az első lábujj, az úgynevezett hallux rövid volt, csak a másik három érintkezett a talajjal, melyek közül a középső hosszabb volt a többinél.[2] Az Albertosaurus képes lehetett 14–21 km/h-s sebesség elérésére is.[10]
Osztályozás és rendszertan[szerkesztés]
| A Tyrannosauridae kladogramja | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Az Albertosaurus a theropoda (a hüllőmedencéjű dinoszauruszok két lábon járó csoportja) Tyrannosauridae családhoz tartozó Albertosaurinae alcsalád tagja. Legközelebbi rokona az alig valamivel korábban megjelent Gorgosaurus libratus (melyre néha Albertosaurus libratus néven is hivatkoznak; lásd lentebb).[11] Csak erről a két albertosaurina fajról készült leírás, de elképzelhető hogy több is létezett.[12] Thomas R. Holtz, Jr. 2004-ben úgy ítélte meg, hogy az Appalachiosaurus is az Albertosaurinae alcsalád tagja,[2] de egy későbbi, még publikálatlan műve,[13] más szerzőkhöz hasonlóan a Tyrannosauridae családba sorolja be.[14]
A tyrannosauridák másik nagyobb alcsaládjába, a Tyrannosaurinae-be tartozik a Daspletosaurus, a Tarbosaurus és a Tyrannosaurus. Az erőteljes felépítésű tyrannosaurináknál gyengébb albertosaurinák aránylag kis koponyával rendelkeztek, a hátsó lábaik alsó lábszára (sípcsontja) és lábfeje (lábközépcsontjai és lábujjai) pedig hosszabbak voltak.[9][11]
A felfedezések története[szerkesztés]
Az Albertosaurust Henry Fairfield Osborn nevezte el a Tyrannosaurus rexről 1905-ben készült leírásának végén levő rövid megjegyzésben. A név Albertára a kanadai tartományra utal, ahol az első maradványait megtalálták. A névhez a dinoszaurusz nevekre jellemzően az ógörög σαυρος / szaürosz ('gyík') szó kapcsolódik. A típusfaj, az A. sarcophagus nevének jelentése 'húsevő', melynek etimológiája megegyezik a szarkofág szóéval, ami az ógörög σαρξ / szarx ('hús') és Φαγειν / phagein ('eszik') szavakból ered.[15] Eddig több mint harminc, különböző korú példány vált ismertté.[4][12]
Korai felfedezések[szerkesztés]

A típuspéldány egy részleges koponya, amit 1844-ben, Albertában, a Horseshoe Canyon-formációban, a Red Deer folyó mentén, egy külszíni lelőhelyen fedeztek fel. Ezt a példányt és egy kisebb koponyát, néhány csontváz maradvánnyal együtt a Kanadai Geológiai Szolgálat (Geological Survey of Canada) Joseph B. Tyrrell által vezetett expedíciói találták meg. A két koponyát 1892-ben Edward Drinker Cope a korábbi „Laelaps incrassatus” fajhoz társította annak ellenére,[16] hogy a Laelaps nevet lefoglalták egy atkanem számára, és Othniel Charles Marsh 1877-ben Dryptosaurusra változtatta. Cope megtagadta a riválisa által alkotott új név használatát, így a „Laelaps incrassatust” Lawrence Lambe nevezte át Dryptosaurus incrassatusra, mikor 1904-ben leírást készített a maradványokról.[17] Nem sokkal ez után Osborn rájött, hogy a D. incrassatus egy általános tyrannosaurida fogon alapul, így a két albertai koponya nem biztos, hogy a fajhoz tartozik. Emellett az albertai koponyák határozottan megkülönböztethetők a D. aquilunguistől, a Dryptosaurus típusfajától, így Osborn 1905-ben egy új nevet hozott létre a számukra, az Albertosaurus sarcophagust, melyhez a Lambe által készített leírást használta fel.[15] Mindkét példányt (CMN 5600 és 5601 jelzéssel) az ottawai Kanadai Természeti Múzeumban (Canadian Museum of Nature) helyezték el.
A Dry Island-i csontmeder[szerkesztés]
1910-ben, egy amerikai őslénykutató, Barnum Brown, rábukkant egy nagyobb Albertosaurus csoport maradványaira a Red Deer folyó mentén, egy másik lelőhelyen. A csontok nagy száma és az idő rövidsége miatt Brown csapata nem gyűjtötte be valamennyi példányt, de azokat igen, amiket azonosítani tudtak a csontmederben. A New York-i Amerikai Természetrajzi Múzeum (American Museum of Natural History) gyűjteményében elhelyezett csontok között hét jobb lábközépcsont és két olyan különálló lábujj csontjai találhatók, amik méretben nem illenek egyik lábközépcsonthoz sem. Ez arra utal, hogy legalább kilenc egyed volt együtt. 1997-ben a Royal Tyrrell Őslénytani Múzeum (Royal Tyrrell Museum of Palaeontology) kutatói újra felfedezték a csontmedret, és folytatták a feltárást a lelőhelyen, ami napjainkban a Dry Island Buffalo Jump Provincial Park területén található.[18] 1997 és 2005 között újabb 13 különböző korú egyed maradványait találták meg, köztük egy apró két év körüli és egy nagyon idős, körülbelül 10 méter hosszúságú példányét is. Egyikük csontváza sem volt teljes, és a legtöbbjük darabjait a két múzeum között osztották szét.[4][5]
Gorgosaurus libratus[szerkesztés]


1913-ban Charles H. Sternberg egy másik tyrannosaurida csontvázát fedezte fel a valamivel régebbi albertai Dinosaur Park-formációban. Egy évvel később Lawrence Lambe a Gorgosaurus libratus nevet adta a leletnek.[19] Később további példányokra bukkantak Alberta és Montana államok területén. Az alapján, hogy kevés különbség található a két nem között, Dale Russell a Gorgosaurust az Albertosaurus fiatal szinonimájává tette, és mivel az Albertosaurus nevet korábban alkották meg, a G. libratust 1970-ben átnevezte Albertosaurus libratusra. Ezáltal az Albertosaurus nem megjelenésének időpontja több millió évvel korábbra került, az elterjedési területe pedig megnövekedett néhány száz kilométerrel déli irányban.[1]
2003-ban Phil Currie több tyrannosaurida koponya összehasonlítása után arra a következtetésre jutott, hogy a két faj jobban eltér egymástól, mint ahogy korábban gondolták. A döntés, hogy egy vagy két nemet használjanak tetszőleges, mivel a két faj testvértaxont alkot, melyek sokkal közelebb állnak egymáshoz, mint bármelyik másik fajhoz. Ezt felismerve Currie azt javasolta, hogy az Albertosaurus és a Gorgosaurus maradjanak külön nemek, mivel nem hasonlítanak jobban egymásra, mint a Daspletosaurus és a Tyrannosaurus, melyek mindig is elkülönültek. Emellett számos albertosaurina példányt fedeztek fel Alaszkában és Új-Mexikóban, amikkel kapcsolatban Currie úgy vélte, hogy amennyiben teljes leírás készül róluk, segíthetnek tisztázni az Albertosaurus-Gorgosaurus helyzetet.[12] Számos szerző követte Currie ajánlását,[2][5][20] de voltak akik eltértek attól.[14]
Egyéb felfedezések[szerkesztés]

William Parks egy 1928-ban, a Red Deer folyó közelében feltárt részleges csontvázat új fajként írt le, Albertosaurus arctunguis néven,[21] de az 1970-es években úgy ítélték meg, hogy a példány azonos az A. sarcophagussal.[1] Parks (ROM 807 jelzésű) lelete a Royal Ontario Museumban található Toronto-ban. Azóta hat további koponyát és csontvázat fedeztek fel Albertában, melyek különböző kanadai múzeumokba kerültek. Újabb fosszíliákról számoltak be Montana, Új-Mexikó és Wyoming államok területéről, de ezek valószínűleg nem az A. sarcophagushoz, és talán nem is az Albertosaurus nemhez tartoznak.[2][12]
Az Albertosaurus megagracilis egy kis tyrannosaurida csontvázon alapul, amit a montanai Hell Creek-formációban találtak.[22] 1995-ben átnevezték Dinotyrannusra,[23] de a legújabb feltétezés szerint egy fiatal Tyrannosaurus rex példánya.[9]
Ősbiológia[szerkesztés]
Növekedési minta[szerkesztés]

Az Albertosaurus legtöbb korosztálya megtalálható a fosszilis rekordban. A csont hisztológia felhasználásával gyakran meghatározható az egyes állatok kora az elpusztulásuk pillanatában, ami lehetővé teszi a növekedési arányuk meghatározását, és más fajokkal való összehasonlítását. A legfiatalabb ismert Albertosaurus, amit a Dry Island csontmederben találtak, kétéves volt, a hossza 2 méter, a tömege pedig 50 kilogramm körül lehetett. Ugyanezen a lelőhelyen egy 10 méteres példányt is találtak, amely a legnagyobb, ami ismertté vált, és egyben a legidősebb is; 28 éves lehetett. Mikor egy közepes korú és tömegű példány adatait egy növekedési grafikonon ábrázolják, az eredmény egy S alakú görbeként jelenik meg, ami szerint a növekedés egy négyéves időszakban a leggyorsabb, ami a tizenhetedik életév körül végződik, a többi tyrannosauridára jellemző módon. A növekedési arány ebben a fázisban 122 kilogramm volt évente, egy 1,3 tonnás felnőttet feltételezve. Más vizsgálatok ennél nagyobb tömeget valószínűsítenek a felnőttek esetében, ami kihat a növekedési arány nagyságára, a növekedési mintára azonban nem. Az Albertosaurushoz hasonló méretű tyrannosauridák növekedési aránya hasonló volt, habár a jóval nagyobb Tyrannosaurus rex a leggyorsabb fázisban ötször gyorsabban gyarapodott (évente 601 kilogrammal).[4] A gyors növekedési fázis vége a feltételezés szerint egybeesik az Albertosaurus ivaréretté válásával, a növekedés azonban kisebb ütemben tovább folytatódott az állat élete végéig.[4][5] Az ivarérettség növekedési szakaszban történő kialakulása egyaránt jellemző a kis[24] és a nagy méretű[25] dinoszauruszokra, ahogy a nagyobb emlősökre, például az emberre vagy az elefántra is.[25] Ez a jellegzetesség jelentős eltérést mutat attól, ami a madaraknál tapasztalható, melyeknél az ivarérettség csak a növekedés befejeződése után alakul ki.[20][25]
Élettörténet[szerkesztés]

A legtöbb ismertté vált Albertosaurus egyed 14 éves vagy annál idősebb volt, amikor elpusztult. A fiatal állatok több okból is ritkán találhatók meg kövület formájában, ugyanis kevésbé valószínű, hogy a felnőttekénél jóval kisebb csontjaik fosszilizálódnak, és a fosszíliagyűjtők is kisebb eséllyel találnak rájuk.[26] A fiatal Albertosaurusok aránylag nagyok egy fiatal állathoz képest, a maradványaik mégis ritkábban kerülnek elő a felnőttekénél. A feltételezés szerint ez a jelenség inkább az élettörténetük következménye, mint a fosszilis rekord hiányossága, ugyanis a fiatal Albertosaurusok egyszerűen ritkábban pusztultak el, mint a felnőttek.[4]
Az Albertosaurus élettörténetével kapcsolatos egyik elképzelés szerint a fiókák nagy számban hullottak el, de kis méretük és törékeny felépítésük miatt nem fosszilizálódtak. Kétéves kor felett a fiatalok már a felnőttekhez hasonlóan nagyobbak voltak a területükön élő összes többi ragadozónál, a zsákmányaiknál pedig gyorsabban futottak. Ez drasztikus módon lecsökkentette a pusztulási arányukat, ami összhangban áll a fosszíliák hiányával. A pusztulási arány 12 éves korra kétszeresére növekedett, feltehetően a felgyorsult növekedés követelményei folytán, 14 és 17 éves kor között, az ivarérettség állapotának elértével pedig ismét megduplázódott. Az elhullás mértéke tovább növekedett a felnőttkor során, feltehetően a magas fizikai követelmények, a stressz és a sérülések miatt, amik részben a fajtársakkal az erőforrásokért és a párzásra alkalmas egyedekért vívott összetűzésekből, továbbá az öregedés egyre növekvő káros hatásaiból származtak. A felnőttek magasabb pusztulási aránya magyarázattal szolgálhat a kövületeik gyakoribb előfordulására. Az igen nagy méretű állatok viszont ritkák, mivel csak kevés egyed élt elég sokáig ahhoz, hogy ilyen méretet érjen el. A fiókák körében magas pusztulási arány, ami lecsökkent a fiatalok között, majd hirtelen megemelkedett a nemi érettség kialakulásával, nagyon kevés állat számára tette lehetővé a maximális testméret elérését, ahogyan az számos ma élő nagy testű emlős, például az elefánt, a kafferbivaly és az orrszarvú esetében is megfigyelhető. Ugyanez a minta jellemző a többi tyrannosauridára is. A modern állatokkal és más tyrannosauridákkal való összehasonlítás megerősíti ezt az élettörténet elméletet, de a fosszilis rekordban tapasztalható eltérések is nagy szerepet játszhatnak, ugyanis az Albertosaurus példányok kétharmada egyetlen területről származik.[4][20]
Csapatos viselkedés[szerkesztés]
A Barnum Brown és csapata által felfedezett Dry Island csontmeder 22 Albertosaurus maradványait tartalmazta, melyek létszáma meghaladja az együtt talált más kréta időszaki theropodák számát, és a második helyen áll az együtt talált nagy theropodákat illetően a Utah államban, a Cleveland Lloyd dinoszaurusz-lelőhelynél (Cleveland Lloyd Dinosaur Quarry) talált Allosaurus maradványai mögött. Úgy tűnik, hogy az egyik példány nagyon idős lehetett, volt köztük nyolc 17 és 23 év közötti felnőtt, hét, a gyors növekedési fázisban levő 12–16 éves fiatal, valamint hat 2–11 év közötti kisebb egyed, melyek még a növekedési fázis előtt álltak.[4]
A növényevők maradványainak majdnem teljes hiánya, és a csontmederben talált Albertosaurus kövületek állapotának hasonlósága alapján Phil Currie arra következtetett, hogy a hely nem egy olyan ragadozó csapda lehetett, mint a Kaliforniában levő La Brea-i kátránygödrök (La Brea Tar Pits), és hogy az állatok egyszerre pusztultak el. Currie szerint ez bizonyítékkal szolgál a csapatos viselkedésre.[18] Más tudósok azonban szkeptikusak, szerintük az állatok az aszály, az árvíz vagy más ok miatt gyűltek össze.[4][26][27]
Bőséges bizonyíték áll rendelkezésre a növényevő dinoszauruszok társaságkedvelő viselkedésére a ceratopsiák és a hadrosauridák esetében,[28] a ragadozók közül azonban csak ritkán található meg ennyi egy helyen. A kis méretű theropodák, például a Deinonychus,[29] a Coelophysis és a Megapnosaurus (Syntarsus) rhodesiensis[30] gyakrabban találhatók meg felhalmozódva, mint az Allosaurushoz és a Mapusaurushoz hasonló nagyobb méretűek.[31] Létezik bizonyíték más tyrannosauridák csapatos viselkedésére is. Kisebb egyedek töredékes maradványaira találtak a chicagói Field Természetrajzi Múzeumban (Field Museum of Natural History) kiállított „Sue”-nak elnevezett Tyrannosaurus közelében, és a montanai Two Medicine-formáció egyik csontmedrében, ahonnan három Daspletosaurus került elő, számos hadrosaurida társaságában.[32] Ezek a leletek összhangban állhatnak az Albertosaurus társas viselkedésére utaló bizonyítékokkal, bár elképzelhető, hogy némelyikük vagy valamennyiük csupán ideiglenesen vagy természetellenes módon gyűlt össze.[18] Vannak, akik azt feltételezik, hogy az ilyen fosszíliák arra utalnak, hogy ezek az állatok a komodói sárkányhoz hasonlóan közösen táplálkoztak, az egyedek egy része pedig az agresszív versengés következtében a társai áldozatává és táplálékává vált.[26]
Currie felvetette annak lehetőségét is, hogy az Albertosaurus csapatban vadászott. A kisebb példányok lábainak arányai összevethetők a leggyorsabb dinoszauruszok közé tartozó ornithomimosaurusokéval. A fiatalabb Albertosaurusok ugyanolyan gyorsak vagy még gyorsabbak lehettek, mint a zsákmányaik. Currie azt feltételezte, hogy a falka fiatalabb tagjai voltak felelősek a zsákmány idősebb állatok felé tereléséért, melyek jóval erősebbek, de ugyanakkor lassabbak is voltak.[18] A fiatalok talán más életstílust követtek, mint a felnőttek, betöltve az ökológiai fülkéket az óriási felnőttek és a kisebb, velük egyidőben élt theropodák közt, melyek közül a legnagyobbak tömege is két nagyságrenddel elmaradt a felnőtt Albertosaurusokénál.[2] Hasonló a helyzet a mai komodói sárkányoknál, ahol a fiókák még apró rovarokkal táplálkoznak, mielőtt felnőve a sziget domináns ragadozóivá válnának.[33] A viselkedésre utaló bizonyítékok azonban rendkívül ritkák a fosszilis rekordban, így ezek az elméletek nehezen tesztelhetők.
Ősökológia[szerkesztés]

Az Albertosaurus sarcophagus valamennyi azonosítható fosszíliája az albertai Horseshoe Canyon-formációból került elő. Ez a geológiai képződmény, 73–70 millió évvel ezelőttre, a késő kréta időszak kora maastrichti korszakára tehető. Közvetlenül alatta található a tengeri eredetű Bearpaw-formáció, amely a Nyugati Belső Víziút részét képezte. A víziút az éghajlat lehűlésével és a tengerszint csökkenésével a kréta időszak vége felé visszahúzódott, felfedve a korábban víz alatt levő területet. A folyamat nem volt egyenletes, és a tengeri út időszakonként többször újra elborította a Horseshoe Canyont, mielőtt végleg eltűnt volna. A tengerszint változásai miatt számos különböző környezet alakult ki a Horseshoe Canyon-formációban, beleértve a parttól távoli, partközeli és a partmenti élőhelyeket, mint a lagúnák, tölcsértorkolatok és árapálysíkságok. Számos széndarab utal a tőzegmocsarak egykori jelenlétére. A formációban talált legtöbb gerinces fosszíliájához hasonlóan az Albertosaurus maradványai is a nagy folyók deltáiból és az ártéri területekről származnak, a Horseshoe Canyon időszakának második feléből.[34]
A Horseshoe Canyon-formáció faunája jól ismert a gerinces fosszíliák által, melyek között elég gyakoriak a dinoszauruszok maradványai is. A cápák, ráják, tokfélék, iszaphalak, kajmánhalfélék és a kajmánhal-szerű Aspidorhynchus alkották a környék halpopulációját. Az emlősök között megtalálhatóak voltak a Multituberculata csoport tagjai és az erszényes Didelphodonok. A sósvízi plezioszauruszok közé tartozó Leurospondylust is megtalálták a Horseshoe Canyon tengeri üledékeiben, de előkerültek édesvízi állatok, például teknősök, a Champsosaurus valamint a krokodilokat képviselő Leidyosuchus és Stangerochampsa maradványai is. Az állatvilágot a dinoszauruszok uralták, főként az olyan hadrosauridák, mint az Edmontosaurus, a Saurolophus és a Hypacrosaurus, melyek az ismertté vált dinoszauruszok felét alkotják. A ceratopsiák és az ornithomimosaurusok szintén nagyon gyakoriak voltak, belőlük állt a populáció további harmada. A jóval ritkább ankylosaurusokkal és pachycephalosaurusokkal együtt, ezek az állatok szolgáltak a különböző húsevő theropodák, a troodontidák, a dromaeosauridák és a caenagnathidák zsákmányául. A terület csúcsragadozói a felnőtt Albertosaurusok voltak, a közepes méretű ragadozók ökológiai fülkéjét betöltő utódaikkal együtt.[34]
Jegyzetek[szerkesztés]
- ↑ a b c Russell, Dale A. (1970). „Tyrannosaurs from the Late Cretaceous of western Canada”. National Museum of Natural Sciences Publications in Paleontology 1, 1–34. o.
- ↑ a b c d e f g Holtz, Thomas R..szerk.: Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka: Tyrannosauroidea, The Dinosauria, Second Edition, Berkeley: University of California Press, 111–136. o. (2004). ISBN 0-520-24209-2
- ↑ a b Paul, G.S.. The Princeton Field Guide to Dinosaurs, Second, Princeton University Press, 108–109, 113. o. (2016). ISBN 978-0-691-16766-4
- ↑ a b c d e f g h i Erickson, Gregory M., Currie, Philip. J.; Inouye, Brian D.; & Wynn, Alice A. (2006). „Tyrannosaur life tables: an example of nonavian dinosaur population biology”. Science 313, 213–217. o. DOI:10.1126/science.1125721. PMID 16840697.
- ↑ a b c d e Erickson, Gregory M., Makovicky, Peter J.; Currie, Philip J.; Norell, Mark A.; Yerby, Scott A.; & Brochu, Christopher A. (2004). „Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs”. Nature 430, 772–775. o. DOI:10.1038/nature02699.
- ↑ Christiansen, Per, & Fariña, Richard A. (2004). „Mass prediction in theropod dinosaurs”. Historical Biology 16 (2-4), 85–92. o. DOI:10.1080/08912960412331284313.
- ↑ (2014) „Body mass estimation in non-avian bipeds using a theoretical conversion to quadruped stylopodial proportions”. Methods in Ecology and Evolution 5 (9), 913–923. o. DOI:10.1111/2041-210X.12226.
- ↑ (2014) „Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage”. PLoS Biology 12 (5), e1001853. o. DOI:10.1371/journal.pbio.1001853. PMID 24802911.
- ↑ a b c Currie, Philip J. (2003). „Allometric growth in tyrannosaurids (Dinosauria: Theropoda) from the Upper Cretaceous of North America and Asia”. Canadian Journal of Earth Sciences 40 (4), 651–665. o. DOI:10.1139/e02-083. (Hozzáférés: 2008. november 7.)
- ↑ a b Dodson, Peter, Britt, Brooks & Carpenter, Kenneth & Forster, Catherine A. & Gillette, David D. & Norell, Mark A. & Olshevsky, George & Parrish, J. Michael & Weishampel, David B.. Albertosaurus, The Age of Dinosaurs. Publications International, LTD., 106–107. o. (1993). ISBN 0-7853-0443-6
- ↑ a b Currie, Philip J., Hurum, Jørn H; & Sabath, Karol. (2003). „Skull structure and evolution in tyrannosaurid phylogeny” (PDF). Acta Palaeontologica Polonica volume=48 (2), 227–234. o. (Hozzáférés: 2008. november 7.)
- ↑ a b c d Currie, Philip J. (2003). „Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta” (PDF). Acta Palaeontologica Polonica 48 (2), 191–226. o. (Hozzáférés: 2008. november 7.)
- ↑ Holtz, Thomas R.: RE: Burpee Conference (LONG), 2008. november 7. [2016. április 12-i dátummal az eredetiből archiválva]. (Hozzáférés: 2007. június 18.)
- ↑ a b Carr, Thomas D., Williamson, Thomas E.; & Schwimmer, David R. (2005). „A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama”. Journal of Vertebrate Paleontology 25 (1), 119–143. o. DOI:[0119:ANGASO2.0.CO;2 10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2].
- ↑ a b Osborn, Henry F. (1905). „Tyrannosaurus and other Cretaceous carnivorous dinosaurs”. Bulletin of the American Museum of Natural History 21, 259–265. o. (Hozzáférés: 2008. november 7.)
- ↑ Cope, Edward D. (1892). „On the skull of the dinosaurian Laelaps incrassatus Cope”. American Philosophical Society, Proceedings 30, 240–245. o.
- ↑ Lambe, Lawrence M. (1904). „On Dryptosaurus incrassatus (Cope) from the Edmonton Series of the Northwest Territory”. Contributions to Canadian Palaeontology 3, 1–27. o.
- ↑ a b c d Currie, Philip J. (1998). „Possible evidence of gregarious behavior in tyrannosaurids” (PDF). Gaia 15, 271–277. o. [2009. március 26-i dátummal az eredetiből archiválva]. (Hozzáférés: 2008. november 7.) (not printed until 2000)
- ↑ Lambe, Lawrence M. (1914). „On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon”. Ottawa Naturalist 28, 13–20. o.
- ↑ a b c Ricklefs, Robert E. (2007). „Tyrannosaur ageing”. Biology Letters 3 (2), 214–217. o. DOI:10.1098/rsbl.2006.0597.
- ↑ Parks, William A. (1928). „Albertosaurus arctunguis, a new species of therapodous dinosaur from the Edmonton Formation of Alberta”. University of Toronto Studies, Geological Series 25, 1–42. o.
- ↑ Paul, Gregory S.. Predatory Dinosaurs of the World.. New York: Simon & Schuster (1988). ISBN 978-0671619466
- ↑ Olshevsky, George. (1995). „(The origin and evolution of the tyrannosaurids.)” (japán nyelven). Kyoryugaku Saizensen (Dino Frontline) 9, 92–119. o.
- ↑ Erickson, Gregory M., Curry Rogers, Kristi; Varricchio, David J.; Norell, Mark.; & Xu, Xing (2007). „Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition”. Biology Letters 3, 558–561. o. DOI:10.1098/rsbl.2007.0254.
- ↑ a b c Lee, Andrew H., Werning, Sarah (2008). „Sexual maturity in growing dinosaurs does not fit reptilian growth models”. PNAS 105, 582–587. o. DOI:10.1073/pnas.0708903105. PMID 18195356.
- ↑ a b c Roach, Brian T., & Brinkman, Daniel T. (2007). „A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History 48 (1), 103–138. o. DOI:[103:AROCPH2.0.CO;2 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2]. (Hozzáférés: 2008. november 7.)
- ↑ Eberth, David A., & McCrea, Richard T. (2001). „Were large theropods gregarious?”. Journal of Vertebrate Paleontology 21 ((Supplement to 3 - Abstracts of Papers, 61st Annual Meeting of the Society of Vertebrate Paleontology)), 46A. o. (published abstract only)
- ↑ Horner, John R..szerk.: Currie, Philip J.; & Padian, Kevin: Behavior, Encyclopedia of Dinosaurs. San Diego: Academic Press, 45–50. o. (1997). ISBN 0-12-226810-5
- ↑ Maxwell, W. Desmond, Ostrom, John H. (1995). „Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations”. Journal of Vertebrate Paleontology 15 (4), 707–712. o.
- ↑ Raath, Michael A..szerk.: Carpenter, Kenneth & Currie, Philip J.: Morphological variation in small theropods and its meaning in systematics: evidence from Syntarsus rhodesiensis, Dinosaur Systematics: Approaches and Perspectives. Cambridge: Cambridge University Press, 91–105. o. (1990). ISBN 0-521-43810-1
- ↑ Coria, Rodolfo A., & Currie, Philip J. (2006). „A new carcharodontosaurid (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina” (PDF). Geodiversitas 28 (1), 71–118. o. (Hozzáférés: 2008. november 7.)
- ↑ Currie, Philip J., Trexler, David; Koppelhus, Eva B.; Wicks, Kelly; & Murphy, Nate..szerk.: Carpenter, Kenneth: An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA), The Carnivorous Dinosaurs. Bloomington: Indiana University Press, 313–324. o. (2005). ISBN 978-0253345394
- ↑ Auffenberg, Walter. The Behavioral Ecology of the Komodo Monitor. Gainesville: University Press of Florida (2000). ISBN 978-0813006215
- ↑ a b Eberth, David A..szerk.: Currie, Philip J. & Padian, Kevin: Edmonton Group, Encyclopedia of Dinosaurs. San Diego: Academic Press, 199–204. o. (1997). ISBN 0-12-226810-5
Fordítás[szerkesztés]
- Ez a szócikk részben vagy egészben az Albertosaurus című angol Wikipédia-szócikk ezen változatának fordításán alapul. Az eredeti cikk szerkesztőit annak laptörténete sorolja fel. Ez a jelzés csupán a megfogalmazás eredetét és a szerzői jogokat jelzi, nem szolgál a cikkben szereplő információk forrásmegjelöléseként.
További információk[szerkesztés]
- Online Collections Database. [2007. február 24-i dátummal az eredetiből archiválva]. (Hozzáférés: 2008. november 6.)
- List of specimens. [2012. december 27-i dátummal az eredetiből archiválva]. (Hozzáférés: 2008. november 6.)
- Skeletal images. [2007. január 8-i dátummal az eredetiből archiválva]. (Hozzáférés: 2008. november 6.)
- Past lives: Chronicles of Canadian Paleontology. [2006. január 10-i dátummal az eredetiből archiválva]. (Hozzáférés: 2008. november 6.)
- Operation Dino-Lift. [2012. január 29-i dátummal az eredetiből archiválva]. (Hozzáférés: 2008. november 6.)